Apis mellifera

Bal arılarında polen toplamaya yarayan polen sepetçiklerinin oluşması, nektar ve polenle beslenmeye geçiş bu farklılaşmanın en tipik örnekleridir. Hayvanlar aleminin böcekler sınıfında yer alan bal arısının taksonomisi aşağıda verilmiştir.
Apis cinsi içinde “Batı” bal arısı olarak adlandırılan Apis mellifera dışında 3 tür daha bulunur ki bunlar “Doğu” bal arısı türleri olan; Apis cerana, Apis dorsata ve Apis florea’dır. Dünya bal üretiminde A. Cerana’dan kısmen yararlanılırken üretimin tamamına yakın kısmı A. mellifera kullanılarak gerçekleştirilmektedir. Diğer 2 tür ise kovana alınamamış olup doğal yuvalarda tek bir petek üzerinde yaşamaktadırlar.
Arı taksonomisinde türden sonra ırklar yer almaktadır. Örneğin Anadolu ırkı, Apis mellifera anatolica olarak ifade edilir.

Farmakofaj ve farmakofori: bal arısı kolonisinde ( Apis mellifera ) kendi kendine ilaç tedavisi ve hastalık önleme mekanizmaları
- Apiterapi insan halk hekimliğinde hastalıklara çare vaat ediyor ancak bal arısında üretilen ve toplanan bileşiklerin arı sağlığı üzerindeki etkileri daha az biliniyor. Ancak kovan ürünleri, diğer organizmalardan ziyade esas olarak bal arılarının ilaçlanmasını ve sanitasyonunu kolaylaştırmalıdır. Burada hem kendi ürettiği bez salgılarının hem de toplanmış kovan ürünlerinin (farmakognozi) koloni sağlığı üzerindeki etkisini gözden geçiriyoruz. Toplanan bitkilerden elde edilen bileşiklerin antibiyotik aktivitesi çiçek ve bölgesel kökene bağlı olarak oldukça farklılık gösterse de bal, polen ve propolisteki ikincil bitki metabolitleri patojenlere ve parazitlere karşı antibiyotik aktivite açısından önemlidir. Bununla birlikte, arı ürünlerinin belirli arı sağlığını artırıcı aktiviteleri, arıların temel bağışıklık yeterliliğini sağlayan bozulmamış beslenmenin etkilerinden açıkça ayırt edilmelidir. Belirli davranışsal adaptasyonlarla uyum içinde kullanılan aktif madde grupları veya bireysel bileşikler arasındaki etkileşimlerin daha da çözülmesi, bal arılarının koloni sağlığını koruma konusundaki doğal potansiyeline ilişkin anlayışımızı derinleştirecektir.
- Makaleyi tekrar gözden geçir
1. Giriş
Bal arılarının ( Apis mellifera ) koloni temelli yaşam öyküsü ve sosyal organizasyonu, onları herhangi bir parazit veya patojen enfeksiyonu için ideal hedefler haline getirir (Schmid-Hempel 1998 ). Yakın akraba bireylerin yüksek yoğunluğu, sabit çevre koşulları ve yuvadaki zengin protein ve karbonhidrat depoları, büyük omurgalı yırtıcılardan bulaşıcı virüslere kadar uzanan düşmanlar için ideal koşulları sağlar (Mutinelli 2011 ; Schmid-Hempel 1998 ). Bu nedenle, hem bireysel arılara hem de bir bütün olarak koloniye ciddi zarar veren çeşitli parazitik eklembacaklı istilalarının yanı sıra bir dizi bakteriyel, viral, mantar ve protozoan enfeksiyonunun görülmesi şaşırtıcı değildir (Bailey ve Ball 1991 ; Morse ve Flottum 1997) . ; Schmid-Hempel 1998 ). Gerçekten de, bu ajanlardan bazılarının (örn. Varroa destructor , Nosema sp. ve virüsler) yakın zamanda tartışılan küresel koloni kayıplarına dahil olduğu düşünülmektedir (Cornman ve ark. 2012 ; Genersch ve ark. 2010 ; vanEngelsdorp ve ark. 2009 ).
Ancak bu dramatik raporlara rağmen bal arıları, ne bireysel ne de koloni düzeyinde hastalık ve zararlılara karşı hiçbir şekilde savunmasız değildir (Evans ve Spivak 2010). Dış bireysel savunma mekanizmaları (örn. böceğin kütikülü) ve ayrıca sosyal savunma mekanizmaları (örn. yuva boşluğunun tecrit edilmesi), parazitlere ve patojenlere karşı etkili savunma hatları sağlar. Bal arısı genomunun serbest bırakılması, bireysel bağışıklık savunmasının genomik arka planına ilişkin değerli bir fikir verdi (Honeybee Genome Sequencing Consortium 2006 ). Artık dört ana doğuştan gelen bağışıklık yolunun (Toll, Imd, JNK, JAK/STAT) bal arılarının enfeksiyon ve yaralanma üzerine doğuştan gelen bağışıklık tepkisinde rol oynadığını biliyoruz. Patojenleri azaltan veya ortadan kaldıran proteazom bağımlı bozunma, apoptoz, melanizasyon ve antimikrobiyal efektör proteinler dahil olmak üzere en etkili savunma mekanizmaları için geniş bir moleküler araç kutusu kullanılır. Ancak bal arısını diğer böceklerle karşılaştırmak, Apis'in doğuştan gelen bağışıklık sisteminin, bağışıklıkla ilgili genlerin sayısında ciddi oranda azalmaya dayandığını göstermektedir (Evans ve ark. 2006 ). Patojenler için mükemmel koloni içi koşullar ve görünürdeki kontrol ihtiyacı göz önüne alındığında, bu bir sürpriz oldu. Ancak koloni bağışıklığı yalnızca bireysel bağışıklık yeterliliği tarafından değil aynı zamanda dış bağışıklık savunması tarafından da yönetilir (Otti ve ark. 2014 ). Koloni düzeyindeki mekanizmaların da eşit derecede önemli olduğu gösterilmiştir ve bağışıklık genlerindeki azalma, iyi gelişmiş davranışsal savunma mekanizmaları, 'sosyal' veya 'kolektif bağışıklık' tarafından fazlasıyla telafi edilebilir (Cotter ve Kilner 2010 ; Cremer ve diğerleri). 2007 )
Sosyal bağışıklık, yalnızca koloniye herhangi bir parazit alımını azaltmakla kalmayıp aynı zamanda bireyler ve koloniler arasındaki yayılımı ve bulaşmayı da azaltan tüm antiparazitik koloni düzeyindeki mekanizmaları özetler (Cremer ve ark. 2007). Sosyal davranıştan kaynaklanan bu hastalık direnci, Walter Rothenbuhler'in hijyenik davranış üzerine çığır açan çalışmasından beri bilinmektedir (Rothenbuhler 1964 ; Rothenbuhler ve Thompson 1956 ). İşçiler, Paenibacillus larvaları ile enfekte olmuş larvaların bulunduğu hücrelerin kapağını açar ve enfekte olmuş bireyleri koloniden uzaklaştırır. Bu nedenle hijyenik davranış, çeşitli yavru hastalıklarının (örneğin Amerikan yavru çürüklüğü, tebeşir yavruları ve Varroa akarları) kontrolünde seçici yetiştirmede başarıyla kullanılmıştır (Harbo ve Harris 1999 ; Palacio ve diğerleri 2010 ; Rothenbuhler ve Thompson 1956 ; Spivak ve Gilliam 1998 ). . Hijyenik davranış ve sosyal böcek kolonilerinin savunması için tımarlamanın önemi hakkında yeni birkaç inceleme mevcut olduğundan, bu konuyu daha ayrıntılı olarak ele almayacağız ve okuyucuyu Evans ve Spivak'ın ( 2010 ) mükemmel incelemelerine yönlendirmek istiyoruz. Wilson-Rich ve diğerleri. ( 2009 ) ve buradaki referanslar.
Bu incelemeyi, bitki ürünlerini toplayıcı olarak arıların olağanüstü kapasitesinden kaynaklanan davranışsal savunma mekanizmalarına odaklayacağız. Çiçekli bitkiler ve tozlayıcıların birlikte evrimi, bitkileri tozlaşma hizmeti karşılığında nektar ve polen sağlamaya itmiştir. Arıların ilgisini çekebilmesi için çiçekte bulunan nektarın fermente olmaması ve polenin mantar patojenleri ile kontamine olmaması gerekir. Bu nedenle, eğer birlikte evrim bir başarı öyküsü olacaksa, bitkilerin arılara patojen içermeyen yiyecek sağlaması gerekir. Bu nedenle bitkilerin nektara bakteriyel fermantasyonu önleyen antibiyotik ikincil metabolitler eklediğini görmek o kadar da şaşırtıcı değil. Genel olarak bitkilerin kendileri bakteriyel ve fungal patojenlere karşı korunmaya ihtiyaç duyarlar ve oldukça aktif antimikrobiyal ve antifungal bileşiklerden oluşan paketler bilinmekte ve insan tıbbındaki birçok tıbbi tedavide kullanılmaktadır. Tozlaştırıcı arılar (hem sosyal hem de yalnız) bitkilerden polen ve nektar topladıklarında, yalnızca protein ve karbonhidrat aramakla kalmayacaklar; kaçınılmaz olarak, bireysel arı ve koloni üzerinde anında etki yaratacak ve koloni sağlığını iyileştirecek ilişkili ikincil bitki metabolitlerini de besleyeceklerdir. Bal arıları, temel besin ihtiyaçlarını karşılamak için toplayıcılık yaparak kaçınılmaz olarak nektarın veya polenin bir parçası olarak zararlılarla ve patojenlerle mücadelede faydalı olabilecek bileşikleri toplar. Ancak beslenme fonksiyonu ile sağlık fonksiyonu arasında çok önemli bir fark vardır. Gıdanın besin değeri, temel fizyolojik işleyişi sağlayan ve sağlık durumuna bakılmaksızın kolonide her zaman ihtiyaç duyulabilecek karbonhidrat, protein ve diğer temel bileşiklerin miktarıyla ilgilidir. Sağlıkla ilgili bileşiklere hemen ihtiyaç duyulmayabilir ve bu da sorun yaratır. Belirli bir bileşiği üreten bitkiler, belirli hastalıklarla savaşmak için gerektiğinde çiçek açmayabileceğinden, sağlık için özel yiyecek arama mümkün olmayabilir. Dolayısıyla bu bileşiklerin depolanması bir sorun haline geliyor ve bal arısının üstün olduğu yer burasıdır. Çok yıllık kolonilere sahip tümsosyal bal arıları ile diğer tüm tozlaştırıcı arılar arasındaki büyük fark, bal arısı kolonisinin toplanmış bitki ürünlerini uzun süreler boyunca depolama konusundaki olağanüstü kapasitesidir. Bal arısı kolonisine yalnızca arı polen taşıyıcıları içinde değil, aynı zamanda diyetlerinde hayvansal protein gerektiren diğer birçok sosyal böceğe karşı da muazzam bir avantaj sağlayan şey, toplanan çok çeşitli antimikrobiyal maddeleri depolama yeteneğidir. Yiyecek depolama kapasitesi, bal arılarına, kendilerinin veya koloninin sağlık durumuna bağlı olarak, çeşitli depolanmış ürünler arasından uyarlanabilir bir şekilde seçici bir şekilde seçim yapma fırsatı sağlar.
Apiterapinin geniş alanı ve arı ürünlerinin insan tıbbında her zaman ikna edici olmayan kullanımı göz önüne alındığında, antibiyotik bitki sekonder metabolitlerinin etkilerini insan sağlığından ziyade bal arısı sağlığı bağlamında daha geniş bir şekilde incelemek açıkça gecikmiştir (ikincisi daha sonra gözden geçirilmiştir). Crane 1975 ve Ghisalberti 1979 tarafından yapılan bazı eski incelemeler ).
Arılar yerli veya işlenmiş kovan ürünlerini farmakofaji ve farmakofori olarak tanımlanan iki alternatif farmakolojik yolla kullanırlar. Farmakofaj, hastalığı azaltmak veya bal arısı sağlığını artırmak için doğrudan tüketimden (örneğin bal, polen, arı sütü) kaynaklanan tüm savunma mekanizmalarıyla ilgilidir; farmakofori ise yenmeyen kovan ürünlerini (örneğin propolis, reçine) ifade eder (König 1988 ). Toplanan bileşiklere ek olarak, bal arısı tarafından üretilen, doğuştan gelen bağışıklık sistemiyle ilgili olmayan ancak yine de zararlılara ve patojenlere karşı oldukça etkili olan bileşikleri de vurgulayacağız. Bu, özellikle bal arılarının çeşitli salgı sistemlerinde salgılanan bileşikleri içerecektir. Bu nedenle, bal arısı kolonilerinde kendi kendine üretilen ve toplanan bileşikleri ele alacağız ve bu ürünlerin sağlığı geliştirici aktivitelerini tartışacağız.
2 Kendi ürettiği kovan ve arı ürünlerinin antibiyotik ve arı sağlığını iyileştirici faaliyetleri
Bal arılarının bezleri ve glandüler dokuları, tipik olarak çeşitli uçucu ve uçucu olmayan bileşiklerden oluşan çok çeşitli iyi tanımlanmış feromonlar ve diğer salgılar üretir (Mizrahi ve Lensky 1997 ). Hipofaringeal, mandibular, zehir ve balmumu bezlerinin salgılarının, kendi ürettiği arı ürünleriyle kendi kendine ilaç tedavisi açısından en önemli olduğu tespit edilmiştir. Bu ekzokrin bezleri tarafından salgılanan maddeler yalnızca karbonhidratlar, proteinler ve yağ asitleri dahil olmak üzere tüm besleyici birincil elementleri değil aynı zamanda antimikrobiyal aktiviteye sahip bileşikleri de içerir.
2.1 Kütiküler hidrokarbonlar
Bal arısının tüm vücudunu kaplayan kütiküler hidrokarbonlar öncelikle su kaybını önleyen bir bariyer görevi görür (Şekil 1 ). Buna ek olarak, kimyasalların çevreden emilimini de etkilerler, feromon görevi görebilirler ve bal arısı sağlığı açısından en önemlisi, mikroorganizmaların böceğin kütikülüne bağlanmasına veya nüfuz etmesine karşı en etkili engeli oluştururlar (Blomquist ve Jackson 1979 ; Lockey). 1988 ). Kütikül üzerinde, glandüler salgılarla sürekli olarak yenilenen yağlı bir tabaka oluştururlar ve mantar ve bakterilerin arıların vücuduna girmesini engellerler (Blomquist ve ark. 1980 ; Koidsumi 1957 ; Gołębiowski ve ark. 2013 ). Kütiküler hidrokarbon bileşimi bireysel gelişim sırasında değişir ve hem çevreye hem de bireysel sağlık durumuna bağlıdır (Blomquist ve diğerleri 1980 ). Bal arılarının kütiküler mum tabakası temel olarak hidrokarbonlardan, mono ve polyesterden, serbest yağ asitlerinden ve diğer polar maddelerden oluşur (Blomquist ve diğerleri 1980 ). Bu kütiküler hidrokarbonlar, potansiyel olarak içeri giren patojenleri temizleyen bir tür 'vücut losyonu' görevi görür ve bu nedenle bireysel arı sağlığı için çok önemlidir. Ayrıca yağlı hidrokarbonlarda kolayca çözünen bileşikler vücut yüzeyi boyunca taşınabilir. Bu bileşikler, çoğunun antibiyotik aktiviteye sahip olduğu bilinen esterleri ve yağ asitlerini içerir. Bu nedenle, kütiküler hidrokarbonların doğrudan antimikrobiyal veya antifungal işlevi olmamasına rağmen, bunlar kişisel hijyen için çok önemlidir ve mikroorganizmaların yapışmasını önleyen veya onlar için toksik olabilen antibiyotik bileşiklerin vücut yüzeyine yayılması için bir taşıyıcı madde olarak hizmet edebilirler.
Şekil 1. 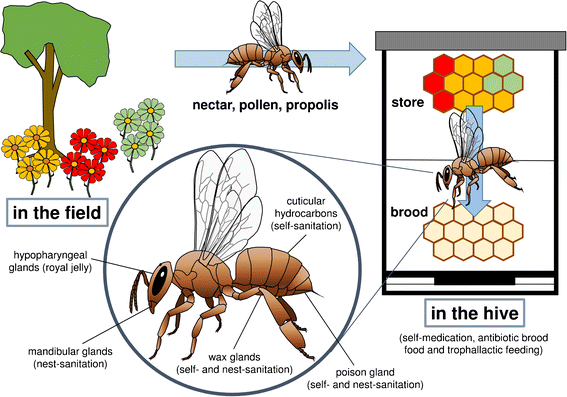
Bal arıları tarafından yalnızca kendilerinin ve yuvaların temizliği için değil, aynı zamanda yavruların ve diğer yuva arkadaşlarının antibiyotikle beslenmesi için de kullanılan, toplanmış ve kendi ürettiği maddelerden oluşan kovan içi repertuar. Tarlada: Antibiyotik potansiyeline sahip ikincil bitki metabolitleri kaçınılmaz olarak polen ve nektarla birlikte toplanır. Yüksek derecede antiviral ve antibiyotik bileşiklere sahip propolis, yuva boşluğunun sanitasyonu için özel toplayıcılar tarafından toplanır. Kovanda: Depolanan bal ve arı ekmeği, hastalıklı ve sağlıklı larvaların ve diğer yuva üyelerinin beslenmesinde seçici olarak kullanılabilir. Arı üzerinde: Antibiyotik aktiviteye sahip glandüler salgılar birey ve koloni sağlığı için kullanılabilir.
2.2 Balmumu
Balmumunun fizyokimyasal ve biyolojik özellikleri merhum Randall Hepburn tarafından yazılan iki monografide kapsamlı bir şekilde açıklanmıştır (Hepburn 1986 ; Hepburn ve diğerleri 2014 ). Balmumu balmumu bezleri tarafından salgılanır ve alkanlar, alkenler, hidrokarbonlar (%14), serbest yağ asitleri (%12), monoesterler (%35), diesterler (%14), hidroksimonoesterler ve bazı küçük bileşenlerin kompleks bir karışımından oluşur. (yağ alkolleri ve hidroksidiesterler) (Hepburn ve diğerleri 2014 ; Mizrahi ve Lensky 1997 ). Balmumunun antibiyotik aktivitesi esterlerden ziyade yağ asitleri için bilinmektedir (Koidsumi 1957 ; Gołębiowski ve diğerleri 2013 ). Balmumu ekstraktlarının arı parazitlerine ve patojenlere karşı biyolojik aktivitesini ele alan yalnızca çok az çalışma vardır, ancak doğal tarak hiçbir zaman biyolojik bir ortamda yerinde test edilmemiştir. Aseton, etanol ve metanol içindeki balmumu ekstraktlarının antimikrobiyal aktiviteye sahip olduğu defalarca kanıtlandı (Tablo I ). İn vitro analizler, Paenibacillus alvei ve P. larvae(Amerikan ve Avrupa yavru çürüklüğü ile ilişkili bakteri) (Lavie 1960a ) dahil olmak üzere bal arısı patojenlerine karşı antibakteriyel aktivitenin yanı sıra, taş yavrularıyla ilişkili Aspergillus flavus , Aspergillus fumigatus ve Aspergillus nigermantarlarına karşı da fungisit aktivitesini doğruladı. hastalığı (Kacániová ve ark. 2012). Her iki analizde de ısıtılan (80 °C) veya ilgili solvent içerisinde 1 saat kaynatılan küçük petek parçalarının ekstraktları kullanıldı. Bu ekstraktlarda inhibitör aktiviteye neden olabilecek herhangi bir potansiyel bileşik tanımlanmamıştır. Üstelik balmumu, yeni inşa edilmiş bakir peteklerden veya yeni salgılanmış balmumu pullarından ziyade eski peteklerden elde edilmişti. Bu nedenle, bu peteklerde yetiştirilen depolanmış bal, polen, reçine veya arı larvalarından aktarılan bileşikler de sadece balmumunun kendisi değil, gözlemlenen antibiyotik etkilere de katkıda bulunmuş olabilir. Bununla birlikte, balmumundaki birçok bileşiğin antibiyotik potansiyeli göz önüne alındığında, hem balmumu üreten bireysel arıların hem de gelişen larvaların balmumundaki antibiyotik potansiyelinden yararlanabileceği büyük olasılıkla görünmektedir.
2.3 Gıda jölesi
Hemşire işçi arılar, larvaları hipofaringeal bezlerin protein açısından zengin bir salgısıyla besler; kraliçe larvalara beslenirse arı sütü adı verilir, işçi larvalara beslenirse işçi jölesi ve erkek arı larvalarına beslenirse erkek arı jölesi adı verilir. Bu gıda jölesi, yetişkin kraliçenin özel diyetidir ancak aynı zamanda tamamlayıcı gıda olarak da olsa koloninin diğer tüm yetişkin üyelerine de beslenir. Gıda jölesi su (%60-70), proteinler (%12-15), şeker (%10-16), lipitler (<%10) ve eser miktarda vitamin, tuz ve serbest amino asitlerden oluşur. Protein fraksiyonunun çoğunluğu, ana arı sütü proteinleri olarak adlandırılan proteinleri içerir (inceleme için bkz. Buttstedt ve ark. 2014 ).
Saf ve hatta suyla seyreltilmiş arı sütü ve işçi sütünün bakterilere ( Enterococcus faecalis ) ve mantarlara ( Ascosphaera apis ve Aspergillus niger ) karşı antibiyotik aktiviteye sahip olduğu gösterilmiştir (Tablo I ). Ek olarak, arı sütü ve işçi sütündeki çeşitli bileşiklerin antimikrobiyal etkileri, P. alvei ve P. larvakültürleri üzerinde asidik, sulu etanol ve eter ekstraktları kullanılarak in vitro test edilmiştir (Tablo I ). Genel olarak arı sütü ekstraktlarının, işçi jölesinden daha yüksek önleyici etkileri vardı (Crailsheim ve Riessberger-Gallé 2001 ). Bitkisel P. larvahücreleri suyla seyreltilmiş arı sütü ile 5 dakikalık tedaviden sonra öldürülürken, sporlar hayatta kalma oranında herhangi bir azalma göstermedi (Hornitzky 1998 ).
Mantar patojeni Aspergillus fumigatus ile yapılan in vivo larva enfeksiyonu çalışmaları , arı sütü konsantrasyonunun artmasıyla larva ölümlerinin azaldığını göstermektedir (Foley ve ark. 2012 ). Bunun diyetteki protein eksikliğinden kaynaklanan yetersiz beslenmeden mi yoksa arı sütündeki antifungal bileşenlerden mi kaynaklandığının belirlenmesi gerekiyor.
İki izole edilmiş protein (ana arı sütü proteini 2 ve defensin-1) ve yağ asidi 10-hidroksi-2-desenoik asit, arı sütünün antimikrobiyal aktivitesinin adayları olarak tartışılmaktadır (Bachanová ve diğerleri 2002 ; Bíliková ve diğerleri. 2001 , 2009 ; Chu ve diğerleri 1992 ; Hornitzky 1998 ). Arı sütünün antimikrobiyal aktivitesi sadece doza bağlı olmayıp aynı zamanda bal arısı kolonileri arasında da önemli ölçüde değişiklik göstermektedir (Rose ve Briggs 1969 ). P. larvalarına dirençli kolonilerden elde edilen arı sütü, duyarlı kolonilerden elde edilenlerden çok daha güçlü antimikrobiyal aktiviteye sahipti (Rose ve Briggs 1969 ). Bal arıları, patojen enfeksiyonlarına yanıt olarak arı sütünün kalitesini ayarlayabilir. P. larvaları ile enfekte olmuş kolonilerde çalışan işçiler, çok daha yüksek miktarlarda antibakteriyel peptitler içeren arı sütü üretirler (Bachanová ve ark. 2002 ). Büyük olasılıkla, arı sağlığını iyileştiren yüksek kaliteli proteinli gıdanın artan antimikrobiyal aktiviteyle birleşimi, koloni içi P. larva titrelerini azaltmak veya ortadan kaldırmak için basit ve etkili bir yöntem sunar. Bununla birlikte, aynı koloni içinde bile arı sütü kalitesinin değişkenliği göz önüne alındığında, beslenme, patojen ve arı sütü kalitesi arasındaki etkileşimlerin etkilerini inceleyen geniş bir açık araştırma alanı olduğu kesindir. Apis mellifera'nınsayısız alt türü veya diğer Asya Apistürleriyle ilgili mevsimsel, koloniye özgü veya biyocoğrafik etkiler hakkında hiçbir şey bilinmemektedir .
Arı sütü, antimikrobiyal ve antifungal etkilerinin yanı sıra parazitlere de müdahale eder. V. yıkıcı akarlarla yapılan arena deneylerinde kullanılan ham ekstraktlar (diklorometan-metanol) ve arı sütü, işçi ve erkek arı jölesi fraksiyonları itici veya durdurucu etkilere sahipti (Tablo I ). Bu akarların üreme döngülerini tamamlamak için neden kraliçe hücreleri yerine erkek arı hücrelerini tercih ettiğini anlamak için gıda jölesi ve V. destructor kullanılarak deneyler yapıldı. Orijinal olarak doğal arı sütü fraksiyonlarıyla karakterize edilen 15 polar yarı kimyasaldan oluşan bir karışımın, V. destructor'a karşı karşılaştırılabilir bir caydırıcı aktiviteye sahip olduğu gösterilmiştir (Drijfhout ve ark. 2005 ). Oktanoik asit, tek kovucu madde olarak laboratuvar ve arazi koşullarında akarlara karşı etkili olan ilk uçucu maddedir (Nazzi ve ark. 2009 ). Bu yağ asidi, arı sütünün kendisi kadar iticidir ve ana arı hücrelerinin iticiliğinde rol oynayabilir (Nazzi ve ark. 2009 ).
Çin sacbrood virüsü (CSBV) ile enfekte olmuş arı larvalarına farklı türde arı sütü (heterospesifik besleme) besleyerek arı sütünün kendi kendini tedavi etme potansiyelini ortaya çıkarmak için tek bir deney yapılmıştır. Arı larvalarının türe özgü olmayan kökenli arı sütü ile beslenmesi olan heterospesifik besleme, Apis ceranalarvalarının Apis mellifera arı sütü ile beslenmesinin, A. cerana larvalarının arı sütü ile beslenmesine kıyasla ölüm oranlarını azalttığını ve dolayısıyla Çin sacbrood virüsüne karşı direnci arttırdığını gösterdi. A. cerana arı sütü. Bu etki Apis mellifera larvalarında gözlenmedi (Zhang ve ark. 2014 ). Heterospesifik besleme, antiviral protein ekspresyonunu indükleyebilir veya arı sütü molekülleri, arı larvalarında Çin kutsal yavru viral replikasyonunu doğrudan engelleyebilir (Zhang ve ark. 2014 ). Sonraki diferansiyel protein ekspresyon analizi, stres tepkisi, fagositoz, antioksidasyon ve enerji metabolizmasında yer alan proteinlerin, heterospesifik beslenmenin ölüm azaltıcı aktivitesini açıklayan adaylar olduğunu buldu (Zhang ve ark. 2014 ).
2.4 Arı zehiri
Bal arısı zehiri, proteinler (fosfolipaz A, hiyalüronidaz), peptitler (melittin, apamin), fizyolojik aktif aminler (histamin, dopamin), şekerler, fosfolipitler ve bazı uçucu bileşiklerin karmaşık bir karışımıdır (Mizrahi ve Lensky 1997 ). Kuru ağırlığının %50'sine kadar olan en aktif bileşik melittindir. Yüksek antibiyotik ve biyolojik aktivitesi, insan apiterapisinde onlarca yıldır araştırılmaktadır (Mizrahi ve Lensky 1997 ; Ratcliffe ve ark. 2011 ). Arı zehrinin bal arısı sağlığını iyileştiren özelliklerini yalnızca birkaç çalışma analiz etti. Dişi bal arıları, kendilerinin ve yuvalarının temizliği için vücut yüzeylerine ve petek balmumuna zehir proteinleri (melittin ve apamin) sürerler (Baracchi ve Turillazzi 2010 ; Baracchi ve diğerleri 2011 ). Bununla birlikte, yüksek antibiyotik potansiyeli göz önüne alındığında, diğer kütiküler bileşiklerle potansiyel etkileşimler de dahil olmak üzere arı zehiri veya zehir proteinlerinin antibiyotik aktivitesi üzerine şaşırtıcı derecede az sayıda çalışma vardır.
3 Toplanan kovan ürünlerinin antibiyotik ve arı sağlığını iyileştirici faaliyetleri
Bal arıları kesinlikle vegan anlamda vejetaryendir: nektar ve polen tek protein ve karbonhidrat besin kaynaklarıdır. Açıkçası, yiyecek arayan işçilerin temel amacı koloni üyelerini beslemek için polen, nektar ve su toplamaktır. İşbölümünün ve işçi uzmanlığının toplayıcıların çeşitli görevlere tahsisini nasıl yönlendirdiği iyi anlaşılmıştır (Sayfa 2013 ). Kovan içi işçiler toplanan polen ve nektarı arı ekmeği ve bala dönüştürür. Her iki ürün de kışlar ve kuraklık gibi kıtlık aşamalarını kapatmak için uzun süre saklanabilir. Ancak polen ve nektar sadece su, şeker ve proteinlerden oluşmuyor. Arı sağlığı ve arı hastalıkları üzerinde önemli etkileri olabilecek eser elementler, vitaminler ve özellikle çeşitli ikincil bitki metabolitleri yer almaktadır (Brodschneider ve Crailsheim 2010 ; Vaudo ve ark. 2015 ).
Ayrıca bitki reçinelerinin toplanması konusunda uzmanlaşmış işçiler de bulunmaktadır. Bu reçineler , yuva boşluğunun potansiyel olarak açıkta kalan bölgelerini kapatmak ve korumak için kullanılan propolis üretmek üzere balmumu ve diğer arı türevi maddelerle karıştırılır (bkz. Bölüm 3.1 ).
3.1 Reçine ve propolis
Reçineler, arılar tarafından toplanan ve propolis adı verilen, bir zamanlar kovanda kullanılan ve bal arıları tarafından polen ve balmumu ile karıştırılan ağaç tomurcuğu, yaprak ve yaradan salgılanan yapışkan maddelerdir. Yuva boşluğunu iyileştirmek ve yeniden şekillendirmek için reçinelerin kullanılması Apis mellifera'ya özgü değildir . İğnesiz arılar (Meliponini) ayrıca reçineyi serumen (balmumu ile karıştırılmış) ve batumen veya geopropolis (balmumu, çamur, tohumlar, tahta veya omurgalı dışkısı ile karıştırılmış) olarak işler (Roubik 2006 ). Propolisin kimyasal bileşimi (karmaşık ve çeşitli karışımlarda 300'den fazla kimyasal bileşik), çiçek kökeni ve doğrulaması, halk hekimliğinde kullanımı ve propolisin genel özellikleri çeşitli incelemelerde kapsamlı bir şekilde özetlenmiştir (Bankova ve diğerleri 2000 , 2014 ; Burdock 1998 ; Ghisalberti 1979) . Greenaway ve diğerleri 1990 ; Marcucci 1995 ; Simone - Finstrom ve Spivak 1987 ; Dişi bitki reçinesinin iki kat daha yüksek aktivitesine sahip cinsel açıdan dimorfik bitkilerde, aynı bitki türlerinin bileşimi bile cinsiyete bağlı olarak belirgin biçimde değişebilir (Lokvam ve Braddock 1999 ).
Bu tür ekstraktlarla (etanol ve metanol) tedavi edilen sağlıklı işçi arılar, bireysel doğuştan gelen bağışıklıklarını azaltır ve detoksifikasyon genlerini yukarı doğru düzenler (Johnson ve diğerleri 2012 ; Simone ve diğerleri 2009 ). Ayrıca farklı etanollü propolis ekstraktlarıyla tedavi edilen sağlıklı kolonilerde çok daha düşük mikrop seviyeleri tespit edildi (Simone ve ark. 2009). Bu etki, ekstraktların ve bakterilerin doğrudan temasından kaynaklanan inhibisyon veya kolonide salınan uçucu bileşiklerle açıklandı (Simone ve ark. 2009 ). Her iki çalışma da toplanmış kovan ürünlerinin kendi kendini tedavi etme potansiyelini iddia eden ilk çalışmalardı.
Reçinenin ve ürünlerinin antibiyotik aktivitesini karakterize eden neredeyse tüm çalışmalarda, çok çeşitli çeşitli solventlerle (aseton, asetonitril, dimetilsülfoksit, etanol, etilasetat, heksan, metanol, petrol eter) veya sadece suyla yapılan ekstraktlar kullanıldı (Tablo II ). İn vitro antibakteriyel aktivite, bal arısı propolisi kullanılarak E. faecalis , P. alvei ve P. larvaları (Amerikan ve Avrupa yavru çürüklüğü ile ilişkili) için doza bağlı bir şekilde iyice doğrulanmıştır (Tablo II ). İklim, yerel çevre ve bitki kökeni, değişen antibiyotik aktivitesinin ana itici güçleridir; ıslak tropik yağmur ormanı tipi iklim propolisi en güçlü aktiviteyi gösterir (Seidel ve ark. 2008 ). Antibiyotik aktivitesi çok stabil görünüyor ve propolisin antibiyotik etkileri zamanla herhangi bir düşüş göstermedi (Schmidt ve ark. 2014 ). Pamuk ağacı ve balsam kavak reçinesi üzerinde yapılan metabolomik araştırmalar, ana bileşik bileşiminde ne önemli mevsimsel ne de bölgesel değişiklikler tespit etti, ancak farklı reçine donör türlerinin P. larvalarınıninhibisyonu açısından farklılık gösterdiği görüldü (Wilson ve ark. 2013 ). Toplayıcı bal arılarının, metabolit içeriğine ve antimikrobiyal aktiviteye bağlı olarak birbiriyle yakından ilişkili birçok reçineli bitki türü arasında ayrı seçimler yaptığını gösterdiler. Bazı çalışmalar, etanolik ve sulu propolis ekstraktlarının püskürtülmesinin veya beslenmesinin de koloni düzeyinde etki gösterebileceğini gösterdi. Tedavi edilen kolonilerin P. larvae enfeksiyonundan kısmen kurtulduğu defalarca gösterilmiştir . Tipik olarak tedavi edilen koloniler, tedavi edilmeyen kontrol kolonileriyle karşılaştırıldığında hastalık baskılaması ve hastalıklı larvalarda azalma gösterdi (Kamel ve diğerleri 2013 ; Lindenfelser 1968 ; Mlagan ve Sulimanovic 1982 ). Ayrıca baldaki P. larva spor yükleri propolis ekstraktı uygulamasından sonra azaldı (Antúnez ve ark. 2008 ). Bu koloni düzeyindeki örnekler, propolis bileşiklerinin ya doğrudan larva bağırsağında bitkisel P. larva hücrelerinin çoğalmasını engellediğini ya da bal arısının bağışıklık sistemini uyardığını göstermektedir (Antúnez ve ark. 2008 ). Propolis bileşiklerinin iki ana grubu olan flavonlar/flavonoller ve flavanonlar/dihidroflavonoller, önerilen mekanizmaları tetikleme olasılığı yüksek adaylardır (Mihai ve ark. 2012 ).
Yakın zamanda yapılan bir çalışma, iklimsel olarak farklı 12 bölgeden çeşitli propolis numunelerini P. larvalarına ve A. apis'e(tebeşir kuluçka patojeni) karşı test etti ve bazı numunelerin, her iki patojene karşı gözlemlenen genel olarak yüksek antibiyotik aktivitesinden saptığını buldu. 12 numuneden dördü, P. larva büyümesinin zayıf inhibitörleriydi, ancak A. apisbüyümesinin iyi inhibitörleriydi ; bu, bu iklim bölgelerindeki reçine mevcudiyetinin, geniş spektrumlu antibiyotik aktivitesi sağlama açısından yetersiz olduğunu ortaya koyuyor (Wilson ve ark. 2015 ). Bu nedenle, çiçek bitki örtüsündeki (reçine donörleri) bölgesel farklılıklar, oldukça değişken terapötik potansiyele sahip antibiyotik aktivitesini çok güçlü bir şekilde belirleyebilir.
Propolis ekstraktları yalnızca A. apis'in değil aynı zamanda A. flavus , Aspergillus fumigatusve Aspergillus niger dahil olmak üzere diğer mantar patojenlerinin büyümesini de engeller (Tablo II ). Tebeşir yavruları ile enfekte olmuş ve V. destructor ile enfekte olmuş kolonilerde reçine toplamanın arttığı gözlemlenmiştir (Simone-Finstrom ve Spivak 2012 ; Popova ve diğerleri 2014 ). Artan miktardaki reçine, kesin nedensel mekanizmalar hakkında herhangi bir bilgi olmaksızın tebeşir yavruları enfeksiyonunun yoğunluğunu azaltmıştır (Simone-Finstrom ve Spivak 2012 ).
Reçinelerin Nosema enfeksiyonları üzerinde de etkisi olduğu öne sürüldü . Apis florea ile yapılan kafes deneylerinde iğnesiz arı serumeninin ( Trigona apicalis ) etanolik ekstraktları Nosema ceranae enfeksiyon oranlarını azaltmıştır (Suwannapong ve ark. 2011 ). Ancak bu çalışmada A. florea'dantoplanan propolis kullanılmamıştır , bu nedenle N. ceranae ile enfekte olmuş bal arılarına ilişkin sonuçların, bir bal arısı kolonisinin kendi kendine ilaç tedavisi bağlamında dikkatle ele alınması gerekir.
Etanol ekstraktlarının sadece patojenik ve parazitik mikroorganizmalar üzerinde değil aynı zamanda daha büyük düşmanlar üzerinde de etkili olduğu görülmektedir (Tablo II ). İn vivo analizlerde balmumu güvesi ( Galleria mellonella ) larvalarına propolis etanolik ekstraktlarının beslenmesi, artan propolis konsantrasyonuyla birlikte balmumu güvesi larva büyümesini, hayatta kalmasını ve ısı üretimini azalttı (Eischen ve Dietz 1987 ; Garedew ve diğerleri 2004a ; Johnson ve diğerleri 1994 ). Bal arısı kolonisinde in vivo olarak propolis ekstraktlarının V. destructor üzerine topikal uygulanması, akar üzerinde narkotik ve öldürücü etkilere neden oldu ve yine larva ısı üretimine olumsuz müdahale etti (Damiani ve ark. 2010 ; Garedew ve ark. 2002 , 2003a ). Aslında Simone-Finstrom ve Spivak ( 2010), koloni düzeyinde olgun dişi akar üretimini azaltmak için propolis tedavisini önerdi.
Çok az çalışma yerli propolisin koloni sağlığı bağlamında nasıl kullanıldığını göstermektedir (Tablo II ). Küçük kovan böceği Aethina tumida'nın istilacı yetişkinleri , yerleşmeyi ve yayılmayı önlemek için 'propolis hapishanelerinde' kapsüllendi (Ellis ve ark. 2003 ; Neumann ve ark. 2001 ). Her iki çalışma da Avrupa ve Güney Afrika burun bal arılarını karşılaştırarak gözlemlenen kapsülleme davranışı ve böceğe karşı etkinlik açısından önemli farklılıkların altını çizdi. Bu mekanizma aynı zamanda koloni içindeki ölü memelileri veya büyük böcekleri izole etmek için de kullanılır (Visscher 1980'de gözden geçirilmiştir ) .
Ham bal arısı propolisi ayrıca bal veya yavru çalan karıncaları ve diğer avcı yırtıcıları yuvadan uzaklaştırmak için kullanılır (Tablo I ). Kovan girişleri, arı kolonilerini korumak için küçük, sürünen böceklere karşı yapışkan bir bariyer olarak propolis tabakasıyla yapıştırılmıştır (Seeley ve ark. 1982 ). Bu tür yapışkan bariyer savunması aynı zamanda mekanik ve kimyasal savunmanın bir kombinasyonu da olabilir. Açık alanda yapılan deneyler, propolisin dokumacı karınca Oecophylla smaragdina'yakarşı itici bir etkisi olduğunu gösterdi . Bu etki bitki reçinelerinin yapışkan ve viskoz özellikleriyle açıklanmıştır (Duangphakdee ve ark. 2009 ).
3.2 Polen ve arı ekmeği
Toplayıcılar tarafından toplanan polen, yaklaşık %50'si serbest amino asitler dahil olmak üzere, ortalama olarak yaklaşık %35 proteinli içeriğe sahiptir. Karbonhidrat miktarları oldukça değişkendir, lipit içeriği %10'un altındadır ve vitaminler, mineraller ve eser elementler %10'un altındaki miktarlarda mevcuttur (Campos ve ark. 2008 ; Mizrahi ve Lensky 1997 ). Protein-karbonhidrat oranları ve kimyasal bileşim sadece çiçek kökenine değil aynı zamanda iklim ve çevre koşullarına, bitki yaşına ve polen gelişimi sırasındaki besin durumuna ve polen tanesinin su içeriğine de bağlıdır (Campos ve ark. 2008 ). Polen arayıcıları, toplanan polen topaklarını doğrudan depolama hücresine bırakır. Depolanan polen daha sonra kusulmuş nektar, bal ve glandüler salgılarla karıştırılmış polenin laktik asit fermantasyonu yoluyla kovan içi arılar tarafından arı ekmeğine dönüştürülür (Brodschneider ve Crailsheim 2010 ). Taze ve işlenmiş polenin besin değeri ve fizikokimyasal özellikleri birbirinden farklı değildir, bu da her ikisinin de arı beslenmesi için eşit derecede önemli olduğunu göstermektedir (Herbert ve Shimanuki 1978). Polen diyeti sadece büyüyen larvalar için değil, aynı zamanda kraliçe arının yumurtlamasını sağlaması ve bakıcı arıların besin jölesi üretmesi için de önemlidir. Diğer yetişkin arıların solunum katsayısı tipik olarak 1'e yakın olmasına ve enerji metabolizmasını karşılamak için öncelikli olarak karbonhidrat kullanmalarına rağmen diyetlerinde proteine de ihtiyaç duyarlar. Bağışıklık ve hastalıklarla mücadele söz konusu olduğunda bu özellikle önemlidir. Alaux ve ark. ( 2010 ), kafesteki bal arısı işçilerinin nutrigenomiklerini ve immün yeterliliğini araştırmış ve polifloral polenle beslenen arılarda monofloral polenle karşılaştırıldığında artan glikoz oksidaz ve fenoloksidaz (PO) aktivitesi bulmuştur. Polen diyeti aynı zamanda polen içermeyen diyetlerle karşılaştırıldığında bal arılarında vitellogenin ve spätzle gen ekspresyonunu ve TOR yolunun ve bağışıklık genlerinin yukarı regülasyonunu arttırdı ve antioksidatif enzimleri arttırdı (Alaux ve ark. 2011 ). Her iki çalışma da polenle beslenmenin temel bağışıklık yeterliliğini etkilediğini ve çiçek kaynaklarının çeşitliliğinin arı sağlığı üzerinde doğrudan etkisi olduğunu vurgulamaktadır. Dolayısıyla besin çeşitliliğindeki kısıtlamalar, bal arılarının etkili bir bağışıklık savunması oluşturamamasına ve dolayısıyla koloni için daha yüksek bir enfeksiyon riskine neden olabilir.
DeGrandi-Hoffmann ve ark. ( 2010 ) polenle beslenmenin deforme kanat virüsü (DWV) enfeksiyonu üzerindeki etkisini kafes deneylerinde test etmiştir. DWV ile enfekte olmuş bal arılarını toz şekerle karıştırılmış taze polenle beslediler. Polenle besleme, kontrollere kıyasla DWV titrelerini azalttı (hiç protein yok). Bunun polenin doğrudan antiviral potansiyelinden mi (DeGrandi-Hoffmann ve ark. 2010 ) yoksa dolaylı olarak arıların bağışıklık sistemini güçlendirmesinden mi kaynaklandığı bilinmemektedir. Yakın zamanda yapılan koloni bazlı bir deney, bal arısı kolonilerinin çeşitli botanik kökenli polenlerle beslenmesinin, akut arı felci virüsü (ABPV), kara kraliçe hücre virüsü (BQCV), DWV ve sacbrood virüsü (SBV) ile düşük enfeksiyon yoğunlukları ile ilişkili olduğunu gösterdi (Antúnez et al. al.2015 ) . Polenin gözlemlenen antiviral aktivitesinin quercetin ve diğer fenolik bileşiklerin varlığından kaynaklandığı düşünülmüştür (Antúnez ve ark. 2015 ). Ancak yine de sonuçlar, yetersiz beslenme veya belirli polen türlerinin potansiyel antiviral aktivitesi ile açıklanabilir. Varroa ve virüs enfeksiyonu kaynaklı bağışıklık değişiklikleri ve (kötü)beslenme ile yapılan ödünleşimler hakkında daha fazla ayrıntı için lütfen DeGrandi-Hoffmann ve Chen'in ( 2015 ) güncel incelemesine bakın.
Polenin antibakteriyel ve antifungal aktiviteleri, etanol, metanol veya su ve su-etanol içindeki solvent ekstraktlarının bal arısı patojen bakterileri ( P. alvei , P. larvae ) ve mantarlar ( A. flavus , Aspergillus fumigatus , Aspergillus ) üzerindeki etkileri test edilerek değerlendirildi. nijer ) (Tablo III ). Smith ve ark. ( 1949 ), sulu polen ekstraktlarının P. larva sporülasyonunu konsantrasyona bağlı bir şekilde inhibe ettiğini gösteren ilk kişi olmuştur . İkinci bir deneyde, bu etkinin eterde suya göre daha fazla çözünen fraksiyonlarla bağlantılı olduğunu gösterdiler ancak ekstraktlardaki spesifik antibiyotik bileşiklerini tanımlamadılar (Smith ve diğerleri 1949 ). Crailsheim ve Riessberger-Gallé ( 2001 ) , doğrudan çiçeklerden toplanan polenlerin, bal arısı korbikulalarından toplanan polen topaklarının ve arı ekmeğinin (su-etanol ekstraktları) P. larvalarına karşı antibakteriyel aktivitesini karşılaştırmıştır . Sürekli olarak konsantrasyona bağlı büyüme inhibisyonu buldular. Dahası, polenin aracılık ettiği büyümeyi önleyici etki, arılar poleni ne kadar çok işlediyse büyük ölçüde arttı. Bu, münhasır olmayan iki mekanizmaya atfedildi: (1) arıların yiyecek bezlerinden veya bal midesinden sıvıları kusarak eklediği maddelerden kaynaklanan artan aktivite veya (2) polenin fermantasyonu, zaten polende bulunan maddeleri aşamalı olarak serbest bırakır (Crailsheim ve Riessberger-Gallé 2001 ).
P. larvaları ile enfekte olmuş kolonilerin polen-su karışımlarıyla beslenmesi , işçi larva ölümlerini azaltmıştır (Rinderer ve ark. 1974 ). Daha yeni kafes deneyleri, polifloral polen ve tatlı özsu balından oluşan bir diyetle beslenen enfekte arılarda protein düzeyinde antimikrobiyal efektörlerin zayıf bir yukarı regüle edildiğini gösterdi (Höcherl ve ark. 2012 ). Yine, ya polen, P. larva ile enfekte olmuş bireylerde bağışıklık sisteminin yukarı regülasyonunu kolaylaştırmış olabilir ya da polenin kısmi sindirimi yoluyla spesifik antibakteriyel bileşenler salınabilir (Rinderer ve ark. 1974). Polifloral polen, monofloral polene kıyasla arı larvalarının kullanıldığı in vivo deneylerde gösterildiği gibi, taş yavrularıyla enfekte olmuş arıların hayatta kalma oranlarını daha da artırır (Foley ve ark. 2012).
Tek bir çalışma haricinde, Nosema apis ve N. ceranae'nin spor gelişimi , çiğ polen veya şurupla karıştırılmış arı ekmeğinin beslenmesiyle engellenememektedir (Tablo III ). Tam tersine, proteinle beslenme yoluyla besin zenginleştirmesi, arıların orta bağırsağını genişleterek Nosema spor üretimindeki artışı kolaylaştırıyor gibi görünüyor (Beutler ve Opfinger 1950 ; Rinderer ve Elliot 1977 ). Bununla birlikte, çeşitli botanik kökenlerden gelen polenlerle beslenen koloniler, yalnızca monofloral polenlerle beslenen kolonilerle karşılaştırıldığında daha düşük N. ceranae spor seviyelerine sahipti (Invernizzi ve ark. 2011 ). Yine, bu sonuç, potansiyel yetersiz beslenme ile gerçek kendi kendine ilaç tedavisi arasındaki ayrım konusunda yetersiz kalıyor.
Artan hayatta kalma oranları ve dolayısıyla daha uzun ömür, mikro parazit enfeksiyonu çalışmalarında (polifloral) polenle beslenmenin genel bir etkisi olabilir. Hem N. apis hem de N. ceranae enfeksiyon kafesi deneyleri, polen beslemesinin kendisi ve özellikle polifloral polen için uzun ömürlülük konusunda karşılaştırılabilir sonuçlar gösterdi (Tablo III ). Çoğunlukla polifloral polen, monofloral polene göre daha iyi sonuçlar elde etmiştir.
Protein ve amino asit kalitesi ve miktarı, polenin sağlığı geliştirici aktivitesine ilişkin ana kriterdir. Örneğin yakın zamanda yapılan bir araştırma, polifloral polenin yüksek protein içeriğine sahip monofloral polen diyetinden mutlaka daha iyi olmadığını gösterdi (Di Pasquale ve ark. 2013 ). Hemşire arıların fizyolojisinin niteliksel ve niceliksel olarak artan amino asit tüketiminden etkilenmesi beklenmektedir. Sonuç olarak, daha kaliteli bir gıda jölesi üretebilirler ve bu da diğer arılarda Nosema enfeksiyonlarının ömrünü kısaltan etkisini ortadan kaldırabilir. Ancak arı kolonileri ve gözlem kovanları üzerinde yapılan deneylerde, her iki yetiştirme yöntemi karşılaştırıldığında tam tersi etkiler bulunmuştur. İlave polen beslemesi, enfeksiyon durumundan bağımsız olarak gözlem kovanlarında arı ömrünü uzatırken, ilave beslemeli enfekte arı kolonilerinde herhangi bir yaşam süresi etkisi gözlenmedi (Mattila ve Otis 2006 ). Arı ömrünün, Nosema enfeksiyonunun ve polen beslemesinin incelenmesi söz konusu olduğunda , kafes, gözlem kovanı ve arı kolonisi deneylerinden elde edilen sonuçlar zıt sonuçlar göstermiştir ve dikkatle değerlendirilmesi gerekmektedir.
Polenli ve polensiz beslenmenin kullanıldığı Varroa akar istilası çalışmaları, istila durumundan bağımsız olarak polene erişimi olan arılar için benzer gen ekspresyon modelleri ( vitellogenin ve bağışıklık gen ekspresyonunda artış, proteolizin yukarı regülasyonu, peptidaz aktivitesi ve karbonhidrat metabolizmasında artış) ortaya çıkardı. Varroa ile istila edilmiş ve istila edilmemiş arılar karşılaştırıldığında , akar istilası çok daha düşük gen ekspresyon seviyelerine ve çoğunlukla hedef genlerin aşağı regülasyonuna yol açtı (Alaux ve ark. 2011 ). Polen beslemesi, Varroa'nın arı metabolizması ve bağışıklığı üzerindeki olumsuz etkilerini tersine çeviremedi (Alaux ve ark. 2011 ). Sonuç olarak Varroaparazitliği, arıların polenin faydalı etkilerine erişmesini engelledi (Alaux ve ark. 2011 ). Koloninin beslenme durumu yalnızca transkriptomik değişiklikleri değil aynı zamanda bireysel arılardaki davranışsal adaptasyonları da etkiledi. Örneğin, Varroa jacobsoni ile istila edilmiş koloniler, daha yüksek polen depolarına sahip olduklarında istilalı yavruların uzaklaştırılmasını arttırmıştır (Janmaat ve Winston 2000 ).
Bal arısı larvaları tipik olarak korbiküler polen yerine arı ekmeği formundaki polenle beslenir (Brodschneider ve Crailsheim 2010). Şu ana kadar arı ekmeğinin neden olduğu faydalı arı sağlığı etkileri yalnızca Nosemaile enfekte olmuş kafesli bal arıları üzerinde çalışıldı (Beutler ve Opfinger 1950 ; Basualdo ve diğerleri 2014 ; Porrini ve diğerleri 2011 ). N. apis ve N. ceranae ile enfekte olmuş işçi arı ekmeğini şeker şurubu ile karıştırılarak veya hammadde olarak beslemek, polen için zaten bilindiği gibi, daha uzun ömürlülüğe yol açar . Fizyolojik düzeyde, N. ceranae ile enfekte olmuş arılar, arı ekmeğiyle beslenirlerse hemolimf protein titrelerini artırır, bu da onların parazit enfeksiyonunu tolere etmelerine yardımcı olabilir (Basualdo ve ark. 2014 ). Poleni koloni sağlığı için potansiyel öneme sahip, toplanmış ve depolanmış bir madde olarak ele aldığımızda, polenin bir besin kaynağı olarak bariz rolü nedeniyle ampirik verilerin yorumlanması belirsizliğini koruyor. Gıda kalitesini spesifik bileşiklerin doğrudan tıbbi etkilerinden ayırmak mümkün olmadığı sürece, potansiyel yetersiz beslenmenin ve bozulmuş bağışıklığın etkilerini, polen türevi antimikrobiyal bileşiklerin eksikliğinden kaynaklanan etkilerden ayırmak en iyi ihtimalle zor olacaktır.
3.3 Bal
Bal, esas olarak ana şeker bileşikleri olarak glikoz ve fruktoz ile sudan (~%20) oluşur. Ayrıca çok daha düşük düzeyde di- ve polisakkaritleri, mineralleri, amino asitleri ve hatta bazı proteinleri de içerir (Crane 1975 ; Doner 1977 ). Yüksek şeker konsantrasyonu, herhangi bir mikrop için ölümcül olan güçlü bir ozmotik basınçla sonuçlanır (simbiyotik laktik asit bakterileri hariç, Olofsson ve Vásquez 2008 ). Bal arılarının balın fermente olmasını önleyebilmesinin temel nedeni budur. Peki, eğer yüksek şeker konsantrasyonu her şeyi hallediyorsa, neden balın diğer antimikrobiyal etkileriyle ilgilenelim ki? Aslında seyreltilmiş bala da bakmak için her türlü neden var. Bal, yüksek şeker konsantrasyonlarıyla saklanmasına rağmen, larva yemine eklendiği anda seyreltilir ve dolayısıyla ozmotik basıncın tek başına antimikrobiyal bir faktör olarak yetersiz olduğu daha düşük konsantrasyonlarda antimikrobiyal etkilere sahip olması durumunda oldukça adaptif olacaktır. Yüksek şeker konsantrasyonu dışında çeşitli antimikrobiyal bileşikler ve kimyasal özellikler tanımlanmıştır (Molan 1992a , b ), bunlar arasında glikoz oksidaz tarafından üretilen hidrojen peroksit, düşük pH (asitlik), metilglioksal, antimikrobiyal peptit arı defensin-1, ana arı sütü proteini 1 yer almaktadır. ve balın antimikrobiyal potansiyeli için önemli olan çeşitli fenolik bileşikler (Brudzynski ve Sjaarda 2015 ; Dustmann 1979 ; Kwakman ve Zaat 2012 ). Ayrıca balda bitkiye özgü pek çok sekonder metabolit tespit edilmiştir.
Çoğu çalışmada doğal (işlenmemiş) bal kullanılmış, bu da ozmoz ve antibiyotik bileşiklerin etkileri arasında ayrım yapmayı çoğunlukla zorlaştırmaktadır (Tablo IV ). Balın bal arısı patojenleri üzerindeki antibakteriyel aktivitesini incelemek için iki çalışmada solvent ekstraktı fraksiyonları (metanol-su ve aseton) kullanıldı. Bu garip bir yaklaşım gibi görünmektedir, çünkü kolonide bileşikler yalnızca su çözücü durumunda çalışacaktır (Tablo IV ). Doğal balı P. alvei ve P. larvalarına karşı test eden 1950'ler-1960'lardaki tarihi Fransız çalışmaları dışında (Gonnet ve Lavie 1960 ; Lavie 1960b ; Verge 1951 ), esas olarak Avrupa yavru çürüklüğü ile ilişkili bakteri türü E. faecalis test edilmiştir ( Tablo IV ). Daha yeni araştırmalar, çeşitli antimikrobiyal maddeler arasındaki etkileşimlerin balın Amerikan ve Avrupa yavru çürüklüğü bakterilerine karşı antibakteriyel aktivitesini nasıl etkilediğini ele aldı. Balın türüne bağlı ve balın patojenine özgü etkileşim etkileri belirgindi (Bobiş ve ark. 2011 , 2013 ; Erler ve ark. 2014). Örneğin, siyah keçiboynuzu balı, Avrupa yavru çürüklüğüne özgü ve Avrupa yavru çürüklüğü ile ilişkili bakterilerin büyümesini ayçiçeği balından çok daha etkili bir şekilde inhibe ederken, ayçiçeği balı, P. larva türlerinin büyümesini siyah keçiboynuzu balından daha güçlü bir şekilde inhibe etmiştir (Erler ve ark. 2014) . ).
Alkaloitler, fenolik asitler (örneğin kafeik, p-kumarik, ellagik ve gallik asit) ve flavonoidler (örneğin krisin, galangin, pinocembrin, kersetin, kaempferol ve tektokrisin) dahil olmak üzere ikincil bitki metabolitleri, gözlemlenen etkileşim özgüllüğüne neden olan başlıca adaylardır (Erler ve ark. .Cushnie ve Lamb 2005 ) . Saf bal (Tablo IV ) ve damıtılmış fraksiyonlar (Obaseiki-Ebor ve diğerleri 1983 ) kullanılarak yapılan in vitro analizler, bu bileşiklerin aynı zamanda taş yavru patojenleri Aspergillus flavus ve Aspergillus niger'e karşı antifungal aktiviteye sahip olduğunu gösterdi .
Nosema spp.'ye odaklanan çeşitli çalışmalar . (Tablo IV ), hareketsiz sporları sayarak balın antibiyotik etkisini ele aldı (Gherman ve diğerleri 2014 ; Gregorc 1993 ; Pohorecka ve Skubida 2004 ). Hem kafes hem de koloni deneyleri, bu tür sporların balda depolandıktan sonra hala bulaşıcı olduğunu gösterdi (ancak bkz. White 1919 ). Spor yükündeki azalmanın doğrudan ölçümü olarak balların antifungal aktivitesi, balın N. apis ve N. ceranae'nin yaşam döngüsünü bir noktada engelleyebileceği varsayımına yol açmaktadır (Tablo IV ). Bununla birlikte bal tüketimi, parazit gelişimini olumsuz etkilese bile her zaman arı sağlığını iyileştirmez. Örneğin, kışlayan bal arılarına verilen çam balı, N. apis gelişimini engelledi; yine de arı ölümlerinin artmasına da neden oldu (Gregorc 1993 ; Pohorecka ve Skubida 2004 ). Tatlı özsu balının çiçek dışı kökeni, çiçek nektarında bulunan sağlığı arttırıcı maddelerden yoksun olabilir.
Depolanan balın farmakofajik etkisi N. apisve N. ceranae enfeksiyonu deneylerinde test edildi. Malone ve ark. ( 2001 ) iki farklı balın (manuka ve kekik balı) etkisini karşılaştırmış ve bunun bireysel arıların N. apis spor yükünü azalttığını bulmuştur. Ancak şeker şurubuyla beslenen arılar en uzun ömürlü olduklarından sonuçlar daha az kesindi (Malone ve ark. 2001 ).
Ayrıca Gherman ve ark. ( 2014 ), N. ceranaeiçin bal türüne özgü spor yükünde azalma olduğunu göstermiştir . Önemli olabilecek çok sayıda münhasır olmayan mekanizma öne sürdüler: Aktif bileşikler (1) Nosema sporlarını öldürebilir, (2) bal arısı bağışıklık sisteminin mikrosporidian enfeksiyonlarına karşı savaşma aktivitesini artırabilir veya (3) Nosema'nın bitkisel formlarının replikasyonunu engelleyebilir. spp. Gerçek mekanizmalar ne olursa olsun, N. ceranaespor yükünün azalmasıyla sonuçlanan bu spesifik bal türleri, aynı zamanda N. ceranaeile enfekte olmuş işçiler tarafından seçilmiş ancak sağlıklı arılar tarafından seçilmemiş, bu da balın kendi kendini tedavi etme potansiyeline işaret ediyor. bireysel arı düzeyinde (Gherman ve ark. 2014 ).
Nutrigenomik çalışmalarda sağlıklı arılardaki transkripsiyonel değişiklikleri ölçmek için balın metanol ve etilasetat ekstraktları kullanıldı (Johnson ve diğerleri 2012 ; Mao ve diğerleri 2011 , 2013 ). Ortak bir sonuç olarak, bu ekstraktlarla beslenen arı gruplarında detoksifikasyon süreçleri ve doğuştan gelen bağışıklık (antimikrobiyal peptit genleri) ile ilgili genler yukarı doğru düzenlenmiştir. Ölçülen transkripsiyonel değişiklikleri tek bir maddenin modüle ettiği görüldü. Sporopollenin monomeri olan fenolik asit p -kumarik asidin (temel bitki sporu ve polen dış duvar bileşiği) bu ekstraktlardan izole edilmiş ve pestisitleri aktif olarak detoksifiye ettiği ve doğuştan gelen bağışıklık sistemi gen ekspresyonunu aktive ettiği gösterilmiştir (Mao ve ark. 2011 , , 2013). p -Kumarik asit balda, polende, arı ekmeğinde ve propoliste tespit edilebilir ancak nektarda tespit edilemez. Nutrasötik ve antibiyotik aktivitesinin kombinasyonu, p -kumarik asitin diğer bal bileşikleriyle (nektardan türetilmiş flavonoidler: pinobanksin, pinobanksin 5-metil eter, pinocembrin) birlikte, doğuştan gelen bağışıklık ve detoksifikasyon için ilave veya sinerjistik sonuçlarla iyi etkileşime girebileceği olasılığını arttırmaktadır. (Mao ve diğerleri 2013 ).
Arı sağlığını geliştiren bitki kökenli ürünlerin ikinci grubu fitohormonlardır. Bitkilerde fizyolojik fonksiyonları düzenleyen bir fitohormon olan absisik asit, nektarda, balda ve bal arılarında her aşamada tespit edilebilmektedir. Bu fitohormon, bireysel işçi bal arılarının ve koloninin bağışıklık tepkisini (hücresel ve humoral) ve yara iyileşmesini artırır (Negri ve ark. 2015 ). Fitohormonlar, bal arıları tarafından çeşitli tıbbi amaçlarla kullanılabilecek yeni bir grup aday maddedir. Şu anda, bu fitohormonların bal arıları için potansiyel işlevini anlamanın henüz başlangıcındayız ve antibiyotik potansiyellerini değerlendirmek için daha fazla çalışmaya ihtiyaç var.
4. Sonuç
Bezle üretilen ve toplanan ürünler, arı parazitleri ve patojenler üzerinde oldukça çeşitli bir spesifikliğe ve etkinliğe sahiptir. Bilim insanları onlarca yıldır bu ürünlerin virüslere, bakterilere ve mantarlara karşı antibiyotik etkilerinin ardındaki moleküler mekanizmaları çözmeye çalışıyor. Bununla birlikte, yüksek çözünürlüklü analitik araçların kullanılabilirliği arttıkça, bulmaca çözülmek yerine giderek daha karmaşık hale geliyor. Giderek azalan tespit limitleri ile yüz binlerce madde tanımlanabilmektedir. Bu bileşikler nasıl etkileşir? Bal arılarının iki tür bal arasında seçim yaparken kullandığı bileşikler bu bileşiklerden hangileridir? Bunlar, modern analitik araçlara rağmen cevaplanması zor sorular olmaya devam ediyor.
Farmakofaji ve farmakofori, kendi kendine üretilen bez ürünleri için yaygın olarak bilinmektedir. Özellikle arı sütünün antimikrobiyal maddeleri (10-hidroksi-2-desenoik asit, majör arı sütü proteini 2 ve defensin-1) arıların yavru hastalıklarına karşı mücadelesinde yardımcı olur. Kendi kendine ilaç tedavisi henüz onlar için kanıtlanmamıştır. Gıda jölesinin varsayılan farmakofajik potansiyelini tam olarak kanıtlamak için mevsimsel ve kovanlar arası antibiyotik değişkenliğini hastalıkla ilişkili beslenme davranışıyla birlikte analiz eden çalışmalara ihtiyaç vardır. Bal arısı patojenlerine karşı antibiyotik aktiviteleri büyük ölçüde bilinmediğinden, balmumu ve arı zehirinin farmakolojik potansiyeli hakkında yalnızca spekülasyon yapabiliriz. Bez salgılarının antibiyotik kökeni, zehir proteinleri ve peptidlerinden bile daha karmaşıktır. Yalnız arılar, mandibular bezden salınan uçucu asiklik terpenoidleri, lipitleri ve yağ asidi türevlerini yuva hücresi sanitasyonu için ve yırtıcı hayvanı kovucu olarak kullanırlar (Cane ve diğerleri 1983 ; Cane 1986 ). Bu fungistatik ve bakteriyostatik maddeler Aspergillus niger ve diğer mikroorganizmalara karşı uygulanır ve ayrıca kendi kendine bakım sırasında da yayılabilir. Proteinli maddeler, lipitler ve asitlerin karışımı potansiyel olarak ilave antibiyotik etkilere sahiptir veya türe özgü hastalıklar üzerinde seçici olarak etki eder.
Tam tersine, yüzlerce çalışmanın sonuçları toplanmış kovan ürünlerinin farmakolojik potansiyeliyle ilgilidir. Kovan içi eczane, kendi kendine ilaç kullanımı için üç ana türde doğal ilaç (bal, polen/arı ekmeği ve propolis) sağlar. Arılar tarafından tüketilemeyen propolis, yalnızca dolaylı kovan ilacı olarak görülebilir ancak yine de çok yüksek bir farmakoforik aktiviteye sahiptir. Bal arıları ve diğer arılar ortak bir hastalık ve yırtıcı hayvan yelpazesini paylaştığından, bitki reçineleri arı toplumlarında yaygın olarak kullanılmaktadır. İğnesiz arıların reçineli ürünlerinin mantar önleyici etkileri olduğu bilinmektedir ( Aspergillus niger ) (Garedew ve diğerleri 2003b ; Muli ve diğerleri 2008 ) ve davetsiz misafirin vücuduna reçine bırakarak A. tumida böceklerini mumyalamak için kullanıldığında oldukça etkilidir (Greco ve diğerleri) Halcroft vediğerleri 2011 ) . Karşılaştırmalı çalışmalar (tek tip reçine ve karışımlar), tek reçinelerin farklı etkilere sahip olabileceğini ve karışımların, yırtıcı karıncalara ve A. tumida'ya karşı kovucu etkiler için fonksiyonel tamamlayıcılığı göstererek daha etkili olduğunu ortaya çıkardı (Drescher ve ark. 2014 ). Farklı bitki türlerinin reçineleri yalnızca farklı organizmaları hedef almaz; aynı zamanda sinerjik olarak da hareket ederler (Drescher ve ark. 2014 ). İğnesiz arılar, karıncalara karşı kovucu etkiyi ve yırtıcı böceklere karşı yapışkan bariyer avantajını birleştirir (Duangphakdee ve diğerleri 2009 ; Schwarz 1948 , Roubik 2006'da gözden geçirilmiştir ). Bir karınca saldırısı sırasında, reçine damlacıklarından bariyerler oluşturmak, karıncaları etkili bir şekilde dolaştırmak (Leonhardt ve Blüthgen 2009 ) ve istilacı karıncaların üzerine 'yapıştırıcı', muhtemelen reçine yayarlar (Lehmberg ve diğerleri 2008 ). Bu örnekler, bal arılarının ve iğnesiz arıların reçineli kovan ürünlerini çok geniş fakat benzer bir farmakoforik şekilde kullandıklarını göstermektedir.
Diğerlerinin yanı sıra flavon/flavonol ve flavanon/dihidroflavonol gruplarını da içeren bitki sekonder metabolitleri, genel olarak gözlemlenen çok yüksek bakterisit ve fungisit aktivitelerinin başlıca adaylarıdır. Reçine ve propolisin antibiyotik aktivitesini açıklayan çalışmaların çoğunda organik çözücü ekstraktları kullanılmaktadır; Arılar yalnızca su topladıkları için bunları hiçbir zaman kullanmayacaklar veya bunlara erişemeyecekler. Sonuç olarak, propolis ekstraktlarının yanı sıra diğer arı ve kovan ürünlerinin ekstraktlarının da incelenmesiyle elde edilen sonuçların, bunların aktivitelerine nedensel açıklamalar sağladığı düşünülerek dikkatle ele alınması gerekir.
Polen ve balın, çeşitli Apis (Tablo III ve IV ) ve Apis olmayan arı türleri için farmakofajik aktivitesinde en yüksek değişkenliğe sahip olduğu gösterilmiştir (Boorn ve ark. 2010 ; Chanchao 2009b ; Chan-Rodríguez ve ark. 2012 ; da Cruz ve ark.). Kimoto - Nira ve Amano 2005 ; Temaru ve diğerleri 2007 ; İğnesiz arı Hypotrigona braunsi için depolanan balın ilaç dışı fakat savunma amaçlı olağanüstü bir kullanımı gözlemlendi . Saha gözlemleri, bal hırsızı Lestrimellita cubeeps'in Hypotrigona kolonilerini istila ettiğini tanımladı . Bir savunma stratejisi olarak işçi arılar koloni girişine bal bırakırlar (Michener 1959 ; de Portekiz-Araújo 1958 ). Bu blokaj balın çalınmasını önledi ve farmakoforinin sadece propolis için değil bal için de mevcut olduğunu gösteriyor.
En azından bal için birçok faktörün balın antibiyotik aktivitesiyle ilişkili olduğu bilinmektedir (hidrojen peroksit, düşük pH, metilglioksal vb.). Bireysel arılar ve yavrular için farmakofaji ve kendi kendine tedavi çalışmalarında giderek daha fazla önem kazanan şey yine ikincil bitki metabolitleridir. Alkaloidler, fenolik asitler (örneğin kafeik, p -kumarik, ellagik ve gallik asit) ve flavonoidler (örneğin krisin, galangin, pinocembrin, kersetin, kaempferol ve tektokrisin) enfekte bireylerde parazit ve patojen yüklerinin azalmasıyla bağlantılıdır. Son çalışmalar, alkaloidlerin, terpenoidlerin ve iridoid glikozitlerin bombus arılarındaki bağırsak hastalığına ( Crithidia bombi ) karşı antimikrobiyal potansiyelini bildirmektedir (Baracchi ve diğerleri 2015 ; Manson ve diğerleri 2010 ; Richardson ve diğerleri 2015). Nektardaki ikincil bitki metabolitleri üzerine yapılan çalışmalar, nektar ve polen toplayan bal ve bombus arıları tarafından paylaşılan oldukça çeşitli yiyecek arama bölgelerinin farmakofajik potansiyelini daha iyi anlamak için tüm arı hastalıkları gruplarını kapsayacak şekilde genişletilmelidir.
Bal arısının doğuştan gelen bağışıklık ve detoksifikasyon sistemini çalıştırmak için genel olarak sağlam bir beslenme gerekli olsa da, belirli salgı bezleri ve toplanan bileşikler, belirli enfeksiyonları önlemek ve bunlarla mücadele etmek için özellikle önemli olabilir. Gelecekteki çalışmalar sadece antibiyotik aktiviteleri ve arıların sağlığı üzerindeki etkileri açısından belirli maddeleri ayırmamalı. Bal arısında kendi kendine ilaç tedavisini kapsamlı bir şekilde anlamak istiyorsak, mükemmel tanıma duyularına sahip bireysel işçi arının, davranışını, zengin glandüler sisteminden gelen potansiyel olarak mevcut bileşiklerle ve kovanda ve çevresinde mevcut olanlarla en üst düzeye çıkarmak için nasıl birleştirdiğini anlamalıyız. kendisinin ve koloninin sağlığı.
Referanslar
Alaux, C., Ducloz, F., Crauser, D., Le Conte, Y. (2010) Diyetin bal arısı bağışıklık yeterliliği üzerindeki etkileri. Biyol. Lett. 6 , 562–565
Alaux, C., Dantec, C., Parrinello, H., Le Conte, Y. (2011) Bal arılarında nutrigenomik: polenin sağlıklı ve varroa ile parazitlenmiş arılar üzerindeki besleyici etkilerinin dijital gen ekspresyonu analizi. BMC Genomik 12 , 496
Antúnez, K., Harriet, J., Gende, L., Maggi, M., Eguaras, M., Zunino, P. (2008) Amerikan Yavru Çürüklüğünün kontrolünde doğal propolis ekstraktının etkinliği. Veteriner. Mikrobiyol. 131 , 324–331
Antúnez, K., Anido, M., Branchiccela, B., Harriet, J., Campa, J., Invernizzi, C., ve diğerleri. (2015) Bal arısı patojenlerinin mevsimsel değişimi ve bunun Uruguay'daki polen çeşitliliği ile ilişkisi. Mikrop. Ekol. 70 , 522–533
Bachanová, K., Klaudiny, J., Kopernický, J., Simúth, J. (2002) Poliakrilamid jel üzerinde bakteriyel büyüme inhibisyonu deneyi yoluyla Paenibacillus larva larvalarına karşı aktif olan bal arısı peptidinin tanımlanması . Apidologie 33 , 259–269
Bailey, L., Ball, BV (1991) Bal Arısı Patolojisi. Akademik Basın Ltd., Londra
Bankova, VS, de Castro, SL, Marcucci, MC (2000) Propolis: kimya ve bitki kökenindeki son gelişmeler. Apidologie 31 , 3–15
Bankova, V., Popova, M., Trusheva, B. (2014) Propolis uçucu bileşikleri: kimyasal çeşitlilik ve biyolojik aktivite: bir inceleme. Kimya Cent. J.8 , 28
Baracchi, D., Turillazzi, S. (2010) Apis mellifera (Hymenoptera: Apidae) bireylerinde MALDI-TOF MS ile belirlenen zehir ve kütiküler peptidlerdeki farklılıklar. J. Böcek Physiol. 56 , 366–375
Baracchi, D., Francese, S., Turillazzi, S. (2011) Yırtıcıya karşı savunmanın ötesinde: bal arısı zehiri, sosyal bağışıklığın bir bileşeni olarak işlev görür. Toxicon 58 , 550–557
Baracchi, D., Brown, MJF, Chittka, L. (2015) Bombus arılarında kendi kendine ilaç tedavisine ilişkin davranışsal kanıtlar? [v2; ref durumu: indekslendi, http://f1000r.es/5ep ] F1000Research 4 , 73
Bastos, EM, Simone, M., Jorge, DM, Soares, AE, Spivak, M. (2008) Brezilya propolisinin Paenibacillus larvalarına karşı antimikrobiyal aktivitesinin in vitro çalışması . J. Omurgasız. Pathol. 97 , 273–281
Basualdo, M., Barragán, S., Antúnez, K. (2014) Arı ekmeği, bal arısı hemolimf proteinini arttırır ve bal arılarında daha yüksek Nosema ceranae bolluğuna neden olmasına rağmen daha iyi hayatta kalmayı destekler. Çevre. Mikrobiyol. Temsilci 6 , 396–400
Beutler, R., Opfinger, E. (1950) Bal arısının ( Apis mellifica ) polenle beslenmesi ve Nosema istilası. Z. Karşılaştır Fizyol. 32 , 383–421
Bíliková, K., Wu, G., Simúth, J. (2001) Potansiyel bir çürüme önleyici faktör olarak bal arısı arı sütünden bir peptid fraksiyonunun izolasyonu. Apidologie 32 , 275–283
Bíliková, K., Mirgorodskaya, E., Bukovská, G., Gobom, J., Lehrach, H., Simúth, J. (2009) Bal arısı arı sütünün azınlık bileşeninin fonksiyonel proteomiklerine doğru: translasyon sonrası değişikliklerin etkisi apalbumin2'nin antimikrobiyal aktivitesi üzerine. Proteomik 9 , 2131–2138
Bíliková, K., Popova, M., Trusheva, B., Bankova, V. (2013) Propolisten saflaştırılan yeni anti- Paenibacillus larva maddeleri. Apidoloji 44 , 278–285
Blomquist, GJ, Jackson, LL (1979) Böcek mumlarının kimyası ve biyokimyası. Prog. Lipid Res. 17 , 319–345
Blomquist, GJ, Chu, AJ, Remaley, S. (1980) Bal arısı Apis mellifera L. Insect Biochem'de balmumu biyosentezi . 10 , 313–321
Bobiş, O., Mărghitaş, LA, Dezmirean, DS, Chirilă, F., Moritz, RFA (2011) Farklı Rumen bal türleri için antioksidan ve antimikrobiyal kapasiteye ilişkin ön çalışmalar. Bülten UASVM Anim. Bilim. Biyoteknoloji. 68 , 91–97
Bobiş, O., Mărghitaş, LA, Dezmirean, DS, Gherman, B., Chirilă, F. (2013) Tek çiçekli balın bal arısı patojenleri Paenibacillus larvae ve Escherichia coli'ye karşı in vitroantibakteriyel aktivitesi . Bilimsel Çalışmalar-Zootehnie Serisi 60 , 139–144
Boonsai, P., Phuwapraisirisan, P., Chanchao, C. (2014) Thai Apis mellifera propolisinden bir kardanolün antibakteriyel aktivitesi . Uluslararası J. Med. Bilim. 11 , 327–336
Boorn, KL, Khor, YY, Sweetman, E., Tan, F., Heard, TA, Hammer, KA (2010) İğnesiz arı Trigona carbonaria'dan elde edilen balın agar difüzyonu, agar seyreltmesi, et suyu mikrodilüsyonu ve zamanla belirlenen antimikrobiyal aktivitesi -öldürme metodolojisi. J. Başvuru. Mikrobiyol. 108 , 1534–1543
Boukraâ, L., Benbarek, H., Ahmed, M. (2008) Diastaz Sayısı ile bağlantılı olarak nişasta ve balın Aspergillus niger'e karşı sinerjistik etkisi. Mikozlar 51 , 520–522
Brodschneider, R., Crailsheim, K. (2010) Bal arılarında beslenme ve sağlık. Apidologie 41 , 278–294
Brudzynski, K., Sjaarda, C. (2015) Antimikrobiyal peptitler, ana arı sütü proteini 1'in jelleinlerini içeren bal glikoproteinleri, balın hücre duvarı litik ve bakterisidal aktivitelerinden sorumludur. PLoS ONE 10 , e0120238
Dulavratotu, GA (1998) Arı propolisinin (propolis) biyolojik özelliklerinin ve toksisitesinin gözden geçirilmesi. Gıda Kimyası Toksikol. 36 , 347–363
Buttstedt, A., Moritz, RFA, Erler, S. (2014) Bal arısının ( Apis mellifera ) başlıca arı sütü proteinlerinin sarı gen ailesinin üyeleri olarak kökeni ve işlevi . Biyol. Rev. 89 , 255–269
Calderone, NW, Lin, S. (2001) Varroa yıkıcının (Acari: Varroidae) işçi ve erkek arı bal arılarının ( Apis mellifera (Hymenoptera: Apidae) larvaları, kozaları ve yavru besinleri ekstraktlarına davranışsal tepkileri). Fizyol. Entomol. 26 , 341–350
Calderone, NW, Lin, S., Kuenen, LPS (2002) Bal arısı, Apis mellifera , işçi ve kraliçe yavrularının parazitik akar Varroa yıkıcıtarafından farklı istilası . Apidologie 33 , 389–398
Campos, MGR, Bogdanov, S., de Almeida-Muradian, LB, Szczesna, T., Mancebo, Y., Frigerio, C., Ferreira, F. (2008) Polen bileşimi ve analitik yöntemlerin standardizasyonu. J. Apic. Res. 47 , 156–163
Cane, JH (1986) Arıların (Hymenoptera: Apoidea) mandibular bez salgılarıyla yırtıcı caydırıcılığı. J. Chem. Ekol. 12 , 1295–1309
Cane, JH, Gerdin, S., Wife, G. (1983) Yalnız arıların (Hymenoptera: Apoidea) çene bezi salgıları: yuva hücresi dezenfeksiyonu potansiyeli. J. Kansas Entomol. Sos. 56 , 199–204
Chanchao, C. (2009a) Tayland'dan Apis dorsata balının özellikleri ve antimikrobiyal aktivitesi . Pak. J. Med. Bilim. 25 , 313–318
Chanchao, C. (2009b) Tayland'dan Trigona laeviceps (iğnesiz arı) balının antimikrobiyal aktivitesi . Pak. J. Med. Bilim. 25 , 364–369
Chan-Rodríguez, D., Ramón-Sierra, J., Lope-Ayora, J., Sauri-Duch, E., Cuevas-Glory, L., Ortiz-Vázquez, E. (2012) Tarafından üretilen balın antibakteriyel özellikleri Gıda kaynaklı mikroorganizmalara karşı Melipona beecheii ve Apis mellifera . Gıda Biyoteknolojisi. 21 , 905–909
Chu, L.-K., Liu, TM-E., Ho, K.-K. (1992) Ascosphaera apis'in arı sütü ve 10-hidroksi-2desenoik asit ile büyümesinin engellenmesi . BI Zool. Acad. Sinica 31 , 73–79
Cornman, RS, Tarpy, DR, Chen, Y., Jeffreys, L., Lopez, D., Pettis, JS, vanEngelsdorp, D., Evans, JD (2012) Çöken bal arısı kolonilerindeki patojen ağları. PLoS ONE 7 , e43562
Cotter, SC, Kilner, RM (2010) Kişisel dokunulmazlığa karşı sosyal dokunulmazlık. Davranış. Ekol. 21 , 663–668
Crailsheim, K., Riessberger-Gallé, U. (2001) Bal arısının Amerikan yavru çürüklüğüne karşı yaşa bağlı direnci. Apidologie 32 , 91–103
Crane, E. (1975) Bal: Kapsamlı Bir Araştırma. Heinemann, Londra
Cremer, S., Armitage, SA, Schmid-Hempel, P. (2007) Sosyal dokunulmazlık. Curr. Biyol. 17 , R693–R702
da Cruz, CBN, Pieri, FA, Carvalho-Zilse, GA, Orlandi, PP, Nunes-Silva, CG, Leomi, L. (2014) İki iğnesiz bal arısı türünden ve Apis mellifera'dan (Hymenoptera: Apidae) elde edilen balların antimikrobiyal aktivitesi patojen mikroorganizmalar. Acta Amaz. 44, 287–290
Cushnie, TPT, Lamb, AJ (2005) Flavonoidlerin antimikrobiyal aktivitesi. Uluslararası J. Antimikrobiyal. Ajanlar 26 , 343–356
Damiani, N., Fernández, NJ, Maldonado, LM, Alvarez, AR, Eguaras, MJ, Marcangeli, JA (2010) Varroa yıkıcısında (Acari: Varroidae) farklı coğrafi kökenlerden propolisin biyoaktivitesi . Parazitol. Res. 107 , 31–37
DeGrandi-Hoffman, G., Chen, Y. (2015) Bal arılarında beslenme, bağışıklık ve viral enfeksiyonlar. Curr. Görüş. Böcek Bilimi. 10, 170–176
DeGrandi-Hoffman, G., Chen, Y., Huang, E., Huang, MH (2010) Diyetin işçi bal arılarında ( Apis mellifera L.) protein konsantrasyonu, hipofaringeal bez gelişimi ve virüs yükü üzerindeki etkisi . J. Böcek Physiol. 56 , 1184–1191
de Portekiz-Araújo, V. (1958) Lestrimelitta cubeps'in (Hymenoptera, Apidae) biyonomiğine katkı . J. Kans. Entomol. Sos. 31 , 203–211
Derevici, A., Popesco, A., Popesco, N. (1964) Propolisin bazı biyolojik özellikleri üzerine araştırmalar. Ann. Arı 7 , 191–200
Di Pasquale, G., Salignon, M., Le Conte, Y., Belzunces, LP, Decourtye, A., Kretzschmar, A., Suchail, S., Brunet, J.-L., Alaux, C. (2013) ) Polen beslenmesinin bal arısı sağlığı üzerindeki etkisi: polen kalitesi ve çeşitliliği önemli mi? PLoS ONE 8 , e72016
Doner, LW (1977) Balın şekerleri-bir inceleme. J. Sci. Gıda Tarımı. 28 , 443–456
Drescher, N., Wallace, HM, Katouli, M., Massaro, CF, Leonhardt, SD (2014) Çeşitlilik önemlidir: arılar farklı reçine kaynaklarından nasıl yararlanır. Ekoloji 176, 943–953
Drijfhout, FP, Kochansky, J., Lin, S., Calderone, NW (2005) Parazitik Varroa akarı Varroayıkıcısının caydırıcıları olarak bal arısı arı sütü bileşenleri . J. Chem. Ekol. 31 , 1747–1764
Duangphakdee, O., Koeniger, N., Deowanish, S., Hepburn, HR, Wongsiri, S. (2009) Bal arılarının ve iğnesiz arıların karınca kovucu reçineleri. Böcekler Soc. 56, 333–339
Dustmann, JH (1979) Balın antibakteriyel etkisi. Apiacta 14 , 7–11
Efem, SEE, Udoh, KT, Iwara, CI (1992) Balın antimikrobiyal spektrumu ve klinik önemi. Enfeksiyon 20 , 227–229
Eischen, FA, Dietz, A. (1987) Bal arısı tarafından toplanan bitki reçinelerini içeren diyetlerle beslenen Galleria mellonella (Lepidoptera: Pyralidae) larvalarının büyümesi ve hayatta kalması. Ann. Entomol. Sos. Am. 80 , 74–77
Ellis, JD, Hepburn, HR, Ellis, AM, Elzen, PJ (2003) Küçük kovan böceğinin ( Aethina tumida Murray) Avrupa bal arıları ( Apis mellifera L.) tarafından sosyal kapsüllenmesi. Böcek Sok. 50 , 286–2
Erkmen, O., Özcan, MM (2008) Türk propolisi, poleni ve defnesinin gıda kaynaklı bozulma ve patojenik mikroorganizmalar üzerindeki antimikrobiyal etkileri. J. Med. Yiyecek. 11 , 587–592
Erler, S., Denner, A., Bobiş, O., Forsgren, E., Moritz, RFA (2014) Bal depolarının çeşitliliği ve bunların bal arısı Apis mellifera'nın patojen bakterileri üzerindeki etkisi . Ekol. Evrim. 4 , 3960–3967
Escuredo, O., Silva, LR, Valentão, P., Seijo, MC, Andrade, PB (2012) Rubus balı değerinin değerlendirilmesi: Polen ve fenolik bileşik içeriği ve antibakteriyel kapasite. Gıda Kimyası 130 , 671–678
Evans, JD, Spivak, M. (2010) Sosyalleştirilmiş tıp: bal arılarında bireysel ve toplumsal hastalık engelleri. J. Omurgasız. Pathol. 103 , S62–72
Evans, JD, Aronstein, K., Chen, YP, Hetru, C., Imler, J.-L., ve diğerleri. (2006) Bal arılarında bağışıklık yolları ve savunma mekanizmaları Apis mellifera . Böcek Mol. Biyol. 15 , 645–656
Fahim, H., Dasti, JI, Ali, I., Ahmed, S., Nadeem, M. (2014) Pakistan'dan gelen Apis dorsata , Apis mellifera ve Ziziphus hünnapbal örneklerinin fiziko-kimyasal analizi ve antimikrobiyal potansiyeli . Asya Pac. J. Trop. Biyomed. 4 , 633–641
Falcão, SI, Vale, N., Cos, P., Gomes, P., Freire, C., Maes, L., Vilas-Boas, M. (2014) Antiprotozoal için Portekiz propolisi ve çiçek kaynaklarının in vitro değerlendirilmesi, antibakteriyel ve antifungal aktivite. Fitother. Res. 28 , 437–443
Foley, K., Fazio, G., Jensen, AB, Hughes, WOH (2012) Bal arısı larvalarında beslenme sınırlaması ve fırsatçı Aspergillusparazitlerine karşı direnç. J. Omurgasız. Pathol. 111 , 68–73
Gallardo-Chacón, JJ, Caselles, M., Izquierdo, M., Rius, N. (2008) Monofloral ve multifloral balların bakteriyel patojenlere karşı inhibitör aktivitesi. J. Apic. Res. 47 , 132–137
Garedew, A., Lamprecht, I., Schmolz, E., Schricker, B. (2002) Propolisin varroacidal etkisi: bir laboratuvar tahlili. Apidologie 33, 41–50
Garedew, A., Schmolz, E., Lamprecht, I. (2003a) Sıcaklığın doğal arı ürünü propolisin antivarroa etkisi üzerindeki etkisinin mikrokalorimetrik ve respirometrik incelenmesi. Thermochim. Acta 399 , 171–180
Garedew, A., Schmolz, E., Lamprecht, I. (2003b) İğnesiz arı Trigona spp.'nin balının antimikrobiyal aktivitesi . J. Apic. Bilim. 47, 37–49
Garedew, A., Schmolz, E., Lamprecht, I. (2004a) Arı tutkalının (propolis), büyük balmumu güvesi Galleria mellonella'nınkalorimetrik olarak ölçülen metabolizma hızı ve metamorfozu üzerindeki etkisi . Thermochim. Acta 413 , 63–72
Garedew, A., Schmolz, E., Lamprecht, I. (2004b) Farklı propolis ekstraktlarının antimikrobiyal etkileri üzerine mikrobiyolojik ve kalorimetrik araştırmalar: in vitro bir yaklaşım. Thermochim. Acta 422 , 115–124
Genersch, E., von der Ohe, W., Kaatz, H., Schroeder, A., Otten, C., ve diğerleri. (2010) Alman arı izleme projesi: Bal arısı kolonilerinin periyodik olarak yüksek kış kayıplarını anlamaya yönelik uzun vadeli bir çalışma. Apidologie 41 , 332–352
Ghaly, MF, Ezzat, SM, Sarhan, MM (1998) Aflatoksijenik mantarları engellemek için propolis ve ultragriseofulvin kullanımı. Folia Mikrobiyol. 43 , 156–160
Gherman, BI, Denner, A., Bobiş, O., Dezmirean, DS, Mărghitaş, LA, Schlüns, H., Moritz, RFA, Erler, S. (2014) Bal arısı Apis mellifera'dapatojenle ilişkili kendi kendine ilaç tedavisi davranışı . Davranış. Ekol. Sosyobiyol. 68 , 1777–1784
Ghisalberti, EL (1979) Propolis: bir inceleme. Arı Dünyası 60 , 59–84
Gołębiowski, M., Cerkowniak, M., Boguś, MI, Włóka, E., Dawgul, M., Kamysz, W., Stepnowski, P. (2013) Calliphora vomitoria'nın kütiküler ve iç lipitlerindeki serbest yağ asitleri ve bunların Antimikrobiyal etkinlik. J. Böcek Physiol. 59 , 416–429
Gonnet, M., Lavie, P. (1960) Balda bulunan antibiyotik faktörü üzerinde ısıtmanın etkisi. Ann. Arı 3 , 349–364
Greco, MK, Hoffmann, D., Dollin, A., Duncan, M., Spooner-Hart, R., Neumann, P. (2010) Alternatif firavun yaklaşımı: iğnesiz arılar, böcek parazitlerini canlı canlı mumyalar. Naturwissenschaften 97 , 319–323
Greenaway, W., Scaysbrook, T., Whatley, FR (1990) Propolisin bileşimi ve bitki kökenleri: Oxford'daki bir çalışma raporu. Arı Dünyası 71 , 107–118
Gregorc, A. (1993) Çam balının bal arısı kolonileri ( Apis mellifera carnica ) ve Nosema hastalığının (nsemosis) gelişimi üzerindeki etkileri . "Prvi Slovenski Veterinarski Kongres, 18–20 Kasım 1993, Portoroz, Slovenya."
Gulfraz, M., Iftikhar, F., Raja, S., Asif, S., Mehmood, S., Anwar, Z., Kaukob, G. (2010) Pakistan'ın çeşitli bal türlerinin kalite değerlendirmesi ve antimikrobiyal aktivitesi. Afr. J. Biyoteknoloji. 9 , 6902–6906
Halcroft, M., Spooner-Hart, M., Neumann, P. (2011) Küçük kovan böceği Aethina tumida'ya karşı iğnesiz arı Austroplebeia australis'in davranışsal savunma stratejileri . Böcekler Soc. 58 , 245–253
Harbo, JR, Harris, JW (1999) Bal arılarında (Hymenoptera: Apidae) Varroa jacobsoni'ye(Mesostigmata: Varroidae) dirençle ilişkili özelliklerin kalıtsallığı. J. Econ. Entomol. 92, 261–265
Hepburn, HR (1986) Bal Arıları ve Balmumu. Springer, Heidelberg
Hepburn, HR, Pirk, CWW, Duangphakdee, O. (2014) Bal Arısı Yuvaları: Kompozisyon, Yapı, İşlev. Springer, Heidelberg
Herbert, EW, Shimanuki, H. (1978) Arılardan toplanan ve arılarda depolanan polenin kimyasal bileşimi ve besin değeri. Apidologie 9 , 33–40
Höcherl, N., Siede, R., Illies, I., Gätschenberger, H., Tautz, J. (2012) Mısırın bal arıları için besin değerinin değerlendirilmesi. J. Böcek Physiol. 58 , 278–285
Bal Arısı Genomu Sıralama Konsorsiyumu (2006) Bal arısı Apis mellifera'nıngenomundan sosyal böceklere ilişkin bilgiler . Doğa 443 , 931–949
Hornitzky, MAZ (1998) Paenibacillus larvaesubsp.'nin patojenitesi . Bal arısı ( Apis mellifera ) kolonilerinde larva sporları ve bitkisel hücrelerin arı sütüne duyarlılığı. J. Apic. Res. 37 , 267–271
Huang, S., Zhang, C.-P., Wang, K., Li, GQ, Hu, F.-L. (2014) Propolisin kimyasal bileşimindeki son gelişmeler. Moleküller 19, 19610–19632
Invernizzi, C., Santos, E., García, E., Daners, G., Di Landro, R., Saadoun, A., Cabrera, C. (2011) Eucaliptus grandis plantasyonlarındaki bal arısı kolonilerinin sıhhi ve beslenme karakterizasyonu . Arch. Zootec. 60 , 1303–1314
Isla, MI, Craig, A., Ordoñez, R., Zampini, C., Sayago, J., Bedascarrasbure, E., Alvarez, A., Salomón, V., Maldonado, L. (2011) Fiziko kimyasal ve biyoaktif Kuzeybatı Arjantin'den gelen balların özellikleri. LWT-Gıda Bilimleri. Teknoloji. 44 , 1922–1930
Janmaat, AF, Winston, ML (2000) Farklı polen depolarına sahip bal arısı kolonilerinde Varroa jacobsoni istilasına uğramış yavruların uzaklaştırılması . Apidologie 31 , 377–385
Johnson, KS, Eischen, FA, Giannasi, DE (1994) Kuzey Amerika arı propolisinin kimyasal bileşimi ve daha büyük balmumu güvesinin ( Lepidoptera : Pyralidae ) larvalarına karşı biyolojik aktivitesi. J. Chem. Ekol. 20 , 1783–1791
Johnson, RM, Mao, W., Pollock, HS, Niu, G., Schuler, MA, Berenbaum, MR (2012) Ekolojik olarak uygun ksenobiyotikler Apis mellifera'da sitokrom P450'leri indükler . PLoS ONE 7 , e31051
Kacániová, M., Vuković, N., Chlebo, R., Hasčík, P., Rovná, K., Cubon, J., Dżugan, M., Pasternakiewicz, A. (2012) Balın, arı poleninin antimikrobiyal aktivitesi Slovakya'dan yükler ve balmumu. Arch. Biyol. Bilim. 64 , 927–934
Kamel, AA, Moustafa, AA, Nafea, EA (2013) Bal arısı kolonilerinde Amerikan yavru çürüklüğü hastalığını kontrol etmek için doğal bir antibiyotik olarak propolis. Afr. J. Agric. Res. 8 , 3047–3062
Kartal, M., Yıldız, S., Kaya, S., Kurucu, S., Topçu, G. (2003) Anadolu'nun iki farklı bölgesinden alınan propolis örneklerinin antimikrobiyal aktivitesi. J. Etnofarmakol. 86 , 69–73
Kimoto-Nira, H., Amano, K. (2008) İğnesiz bal arıları tarafından üretilen balın antimikrobiyal aktivitesi. J. Apic. Res. 47 , 325–327
Koidsumi, K. (1957) Böceklerde kütiküler lipitlerin antifungal etkisi. J. Böcek Physiol. 1 , 40–51
König, B. (1988) Farmakofor böcek olarak bal arısı. Entomol. Jener. 14 , 145–148
Koo, H., Gomes, BP, Rosalen, PL, Ambrosano, GM, Park, YK, Cury, JA (2000) Propolis ve Arnica montana'nın oral patojenlere karşı in vitro antimikrobiyal aktivitesi. Arch. Oral Biyol. 45 , 141–148
Kouidhi, B., Zmantar, T., Bakhrouf, A. (2010) Tunus propolis ekstraktının anti-karyojenik ve anti-biyofilm aktivitesi ve kanser hücrelerinin çoğalmasına karşı potansiyel koruyucu etkisi. Anaerob 16 , 566–571
Kwakman, PHS, Zaat, SAJ (2012) Balın antibakteriyel bileşenleri. IUBMB Hayat 64, 48–55
Lavie, P. (1960a) Arı kolonilerindeki antibakteriyel maddeler ( Apis mellifica L.). Ann. Arı 3 , 103–183
Lavie, P. (1960b) Arı kolonilerindeki antibakteriyel maddeler ( Apis mellifica L.). Ann. Arı 3 , 201–299
Lehmberg, L., Dworschak, K., Blüthgen, N. (2008) İğnesiz arı cinsi Trigona'da (Apidae, Meliponini) karıncalara karşı savunma davranışı ve kimyasal caydırıcılık. J. Apic. Res. 47 , 17–21
Leonhardt, SD, Blüthgen, N. (2009) Yapışkan bir mesele: Bornean'ın iğnesiz arıları tarafından reçine toplanması. Biotropica 41 , 730–736
Lindenfelser, LA (1967) Propolisin antimikrobiyal aktivitesi. Am. Arı J. 107(90–92), 130–131
Lindenfelser, LA (1968) Propolisin Bacillus larvalarına karşı in vivo aktivitesi . J. Omurgasız. Pathol. 12 , 129–131
Lockey, KH (1988) Böcek kütikülünün lipitleri: kökeni, bileşimi ve işlevi. Komp. Biyokimya. Fizik. B89 , 595–645
Logan, A., Ruiz-González, MX, Brown, MJF (2005) Bombus arılarının bağırsak tripanozom parazitinde konakçı açlığının parazit gelişimi ve popülasyon dinamikleri üzerindeki etkisi. Parazitoloji 130 , 637–642
Lokvam, J., Braddock, JF (1999) Clusia grandiflora'nın (Clusiaceae) cinsel açıdan dimorfik tozlayıcı ödüllerinde anti-bakteriyel fonksiyon . Ekoloji 119 , 534–540
Malone, LA, Gatehouse, HS, Tregidga, EL (2001) Bal arısı Apis mellifera'nın(Hymenoptera: Apidae) bir paraziti olan Nosema apis (Microsporidia: Nosematidae) üzerinde zamanın, sıcaklığın ve balın etkileri . J. Omurgasız. Pathol. 77 , 258–268
Manson, JS, Otterstatter, MC, Thomson, JD (2010) Bir nektar alkaloidinin tüketimi, bombus arılarındaki patojen yükünü azaltır. Ekoloji 162 , 81–89
Mao, W., Schuler, MA, Berenbaum, MR (2011) Bal arısındaki ( Apis mellifera ) akarisitlerin CYP9Q aracılı detoksifikasyonu. Proc. Natl. Acad. Bilim. ABD 108 , 12657–12662
Mao, W., Schuler, MA, Berenbaum, MR (2013) Bal bileşenleri, batı bal arısı Apis mellifera'daki detoksifikasyon ve bağışıklık genlerini yukarı doğru düzenler . Proc. Natl. Acad. Bilim. ABD 110 , 8842–8846
Marcucci, MC (1995) Propolis: kimyasal bileşim, biyolojik özellikler ve terapötik aktivite. Apidologie 26 , 83–99
Mattila, HR, Otis, GW (2006) İlkbaharda polen mevcudiyeti ve Nosemaenfeksiyonunun işçi bal arılarının (Hymenoptera: Apidae) iş bölümü ve hayatta kalması üzerindeki etkileri. Çevre. Entomol. 35 , 708–717
Mavri, A., Abramovic, H., Polak, T., Bertoncelj, J., Jamnik, P., Možina, SS, Jeršek, B. (2012) Sloven propolisinin kimyasal özellikleri ve antioksidan ve antimikrobiyal aktiviteleri. Kimya Biyodalgıçlar. 9 , 1545–1558
Michener, CD (1959) Angola'dan Trigona'nın kardeş türleri (Hymenoptera, Apidae). Am. Muş. Novit. 1956 , 1–5
Mihai, CM, Mărghitaş, LA, Dezmirean, DS, Chirilă, F., Moritz, RFA, Schlüns, H. (2012) Propolisin flavonoidleri arasındaki etkileşimler, bal arısı patojeni Paenibacillus larvalarına karşı antibakteriyel aktiviteyi etkiler . J. Omurgasız. Pathol. 110 , 68–72
Mizrahi, A., Lensky, Y. (1997) Arı ürünleri: Özellikler, Uygulamalar ve Apiterapi. Springer, New York, NY
Mlagan, V., Sulimanovic, D. (1982) Propolis solüsyonlarının Bacillus larvaları üzerindeki etkisi . Apiacta 17 , 16–20
Molan, PC (1992a) Balın antibakteriyel aktivitesi - 1. Antibakteriyel aktivitenin doğası. Arı Dünyası 73 , 5–28
Molan, PC (1992b) Balın antibakteriyel aktivitesi - 2. Antibakteriyel aktivitenin gücündeki değişiklikler. Arı Dünyası 73 , 59–76
Morse, RA, Flottum, K. (1997) Bal arısı zararlıları, avcıları ve hastalıkları. Kuzey Arı Kitapları, Medine, Ohio
Muli, EM, Maingi, JM, Macharia, J. (2008) Kenya iğnesiz arısı Dactylurina schimidti'denpropolis ve balın antimikrobiyal özellikleri . Apiacta 43 , 49–61
Mutinelli, F. (2011) Patojenlerin bal arıları ve ürünleri (kraliçe arılar ve sperma dahil) ticareti yoluyla yayılması: genel bakış ve son gelişmeler. Rev. bilim. teknoloji. Kapalı. int. Epiz. 30 , 257–271
Nazzi, F., Bortolomeazzi, R., Della Vedova, G., Del Piccolo, F., Annoscia, D., Milani, N. (2009) Oktanoik asit, arı sütüne varroa önleyici özellikler kazandırır. Naturwissenschaften 96 , 309–314
Negri, P., Maggi, MD, Ramirez, L., De Feudis, L., Szwarski, N., Quintana, S., Eguaras, MJ, Lamattina, L. (2015) Absisik asit Apis mellifera'da bağışıklık tepkisini artırır ve koloni uygunluğuna katkıda bulunur. Apidologie 46 , 542–557
Netíková, L., Bogusch, P., Heneberg, P. (2013) Çek etanol içermeyen propolis özütü, geniş bir yelpazedeki bakteriyel ve fungal patojenlere karşı inhibitör aktivite göstermektedir. J. Gıda Bilimi. 78 , M1421–M1429
Neumann, P., Pirk, CWW, Hepburn, HR, Solbrig, AJ, Ratnieks, FLW, Elzen, PJ, Baxter, JR (2001) Böcek parazitlerinin Cape bal arısı kolonileri ( Apis mellifera capensisEsch.) tarafından sosyal kapsüllenmesi. Naturwissenschaften 88 , 214–216
Amaicha del Valle (Tucuman, Arjantin) propolisinin antibakteriyel aktivitesinin taranması. J. Etnofarmakol. 68 , 97–102
Obaseiki-Ebor, EE, Afonya, TCA, Onyekweli, AO (1983) Bal damıtığının antimikrobiyal aktivitesine ilişkin ön rapor. J. Pharm. Farmakol. 35 , 748–749
Olofsson, TC, Vásquez, A. (2008) Bal arısı Apis mellifera'nın bal midesinde yeni bir laktik asit bakteri florasının tespiti ve tanımlanması . Curr. Mikrobiyol. 57 , 356–363
Otti, O., Tragust, S., Feldhaar, H. (2014) Dış ve iç bağışıklık savunmalarının birleştirilmesi. Trendler Ecol. Evrim. 29 , 625–634
Özkırım, A., Çelemli, Ö.G., Schiesser, A., Charistos, L., Hatjina, F. (2014) Yunan ve Türk propolisinin Paenibacillus larvalarına karşı etkinliklerinin karşılaştırılması . J. Apic. Res. 53 , 528–536
Page Jr., RE (2013) Kovanın Ruhu. Harvard Üniversitesi Yayınları, Cambridge
Palacio, MA, Rodriguez, E., Goncalves, L., Bedascarrasbure, E., Spivak, M. (2010) Deneysel olarak iğneyle öldürülen veya Ascosphaera apis ile enfekte edilen yavrulara yanıt olarak bal arılarının hijyenik davranışları . Apidologie 41 , 602–612
Pohorecka, K., Skubida, P. (2004) Bal arısı kolonilerinin ( Apis mellifera L.) depolarda tatlı özsu balı ilavesiyle kışlamasının sağlığı. Boğa. Veteriner. Öğr. Pulawy 48 , 409–413
Popova, M., Reyes, M., Le Conte, Y., Bankova, V. (2014) Propolisin kimyasal bileşimi ve Varroa yıkıcısına karşı bal arısı direnci . Nat. Ürün. Res. 28 , 788–794
Porrini, MP, Sarlo, EG, Medici, SK, Garrido, PM, Porrini, DP, Damiani, N., Eguaras, MJ (2011) Apis mellifera'da Nosema ceranaegelişimi : diyet ve enfektif aşının etkisi. J. Apic. Res. 50 , 35–41
Radwan, SS, El-Essawy, AA, Sarhan, MM (1984) Balda mikroorganizmalara karşı aktif belirli maddelerin varlığına ilişkin deneysel kanıt. Zentralbl. Mikrobiol. 139 , 249–255
Ratcliffe, NA, Mello, CB, Garcia, ES, Butt, TM, Azambuja, P. (2011) Böcek doğal ürünleri ve süreçleri: insan hastalıkları için yeni tedaviler. Böcek Biyokimyası. Mol. Biyol. 41 , 747–769
Richardson, LL, Adler, LS, Leonard, AS, Andicoechea, J., Regan, KH, Anthony, WE, Manson, JS, Irwin, RE (2015) Çiçek nektarındaki ikincil metabolitler, bombus arılarındaki parazit enfeksiyonlarını azaltır. Proc. R. Soc. B 282 , 20142471
Rinderer, TE, Elliot, KD (1977) İşçi bal arısının Nosema apis enfeksiyonuna tepkisi : diyetin etkisi. J. Econ. Entomol. 70 , 431–433
Rinderer, TE, Rothenbuhler, WC, Gochnauer, TA (1974) Bal arısı larvalarının Bacillus larvalarına duyarlılığı üzerinde polenin etkisi . J. Omurgasız. Pathol. 23 , 347–350
Rose, RI, Briggs, JD (1969) Bal arılarında Amerikan yavru çürüklüğüne karşı direnç IX. Bal arısı larva yeminin Bacillus larvalarının büyümesi ve canlılığı üzerine etkileri. J. Omurgasız. Pathol. 13 , 74–80
Rothenbuhler, WC (1964) Bal arılarında yuva temizliğinin davranış genetiği. I. Dört kendilenmiş soyun hastalık nedeniyle öldürülmüş yavrulara tepkileri. Animasyon. Davranış. 12 , 578–583
Rothenbuhler, WC, Thompson, VC (1956) Bal arılarında Amerikan yavru yavrularına karşı direnç: I. Farklı genetik soylardan larvaların farklı hayatta kalma oranları. J. Econ. Entomol. 49 , 470–475
Roubik, DW (2006) İğnesiz arı yuvalama biyolojisi. Apidologie 37 , 124–143
Şahinler, N., Kurt, S. (2004) Formik asit ve propolis ekstraktının tebeşir hastalığı patojeni Ascosphera apis'e karşı antifungal aktivitesi üzerine bir araştırma . J. Anim. Veteriner. Av. 3 , 554–556
Samšiňáková, A., Kálalová, S., Haragsim, O. (1977) Bazı antimikotiklerin ve dezenfektanların Ascosphaera apis Maassen mantarı üzerindeki in vitro etkileri. J. Başvuru. Entomol. 84 , 225–232
Sauerwald, N., Polster, J., Bengsch, E., Niessen, L., Vogel, RF (1998) Arı sütünün suda çözünür fraksiyonlarının birleşik antibakteriyel ve antifungal özellikleri. Av. Gıda Bilimi 20 , 46–52
Schmidt, EM, Stock, D., Chada, FJG, Finger, D., Sawaya, ACHF, ve diğerleri. (2014) Brezilya taze ve olgun propolisinin karakterizasyonu ve biyolojik özellikleri arasında bir karşılaştırma. BioMed Arş. Uluslararası 2014 , 257617
Schmid-Hempel, P. (1998) Sosyal Böceklerdeki Parazitler. Princeton University Press, Princeton, NJ
Schwarz, HF (1948) Batı yarımkürenin iğnesiz arıları (Meliponidae). B. Am. Muş. Nat. Tarih. 90 , 1–546
Seeley, TD, Seeley, RH, Akratanakul, P. (1982) Tayland'daki bal arılarının koloni savunma stratejileri. Ekol. Monogr. 52 , 43–63
Seidel, V., Peyfoon, E., Watson, DG, Fearnley, J. (2008) Propolisin antibakteriyel aktivitesinin farklı coğrafi ve iklim bölgelerinden karşılaştırmalı çalışması. Fitother. Res. 22 , 1256–1263
Simone-Finstrom, M., Spivak, M. (2010) Propolis ve arı sağlığı: bal arıları tarafından reçine kullanımının doğal tarihi ve önemi. Apidologie 41 , 295–311
Simone-Finstrom, MD, Spivak, M. (2012) Parazit mücadelesinden sonra artan reçine toplama: bal arılarında kendi kendine ilaç tedavisi vakası? PLoS ONE 7 , e34601
Simone, M., Evans, J., Spivak, M. (2009) Bal arılarında reçine toplama ve sosyal bağışıklık. Evrim 63 , 3016–3022
Smith, RL, Beck, JV, Anderson, EJ (1949) Polenlerin Bacillus larvalarının (BEYAZ) sporülasyonu üzerindeki etkisi . J. Bakteriol. 57 , 213–218
Spivak, M., Gilliam, M. (1998) Bal arılarının hijyenik davranışları ve bunun yavru hastalıkları ve Varroa kontrolünde uygulanması. Bölüm I. Hijyenik davranış ve Amerikan yavru çürüklüğüne karşı direnç. Arı Dünyası 79 , 124–134
Stepanović, S., Antić, N., Dakić, I., Svabić-Vlahović, M. (2003) Propolisin in vitro antimikrobiyal aktivitesi ve propolis ile antimikrobiyal ilaçlar arasındaki sinerjizm. Mikrobiyol. Res. 158 , 353–357
Suwannapong, G., Maksong, S., Benbow, ME (2011) İğnesiz arı propolisinin Apis florea'nın Nosema ceranae ile deneysel enfeksiyonu üzerindeki etkileri . J. Agric. Bilim. Teknoloji. A1 , 818–825
Temaru, E., Shimura, S., Amano, K., Karasawa, T. (2007) İğnesiz bal arılarından (Hymenoptera; Apidae; Meliponinae) elde edilen balın antibakteriyel aktivitesi. Pol. J. Microbiol. 56 , 281–285
Tenore, GC, Ritieni, A., Campiglia, P., Novellino, E. (2012) Sicilya siyah bal arıları ( Apis mellifera ssp. sicula ) tarafından üretilen monofloral balların nutrasötik potansiyeli. Gıda Kimyası Toksikol. 50 , 1955–1961
Ulusoy, E., Kolaylı, S., Sarıkaya, AO (2010) Türkiye'den farklı çiçek kökenli balların antioksidan ve antimikrobiyal aktiviteleri. J. Gıda Biyokimyası. 34 , 321–335
Uzel, A., Sorkun, K., Onçağ, O., Coğulu, D., Gençay, O., Salih, B. (2005) Dört farklı Anadolu propolis örneğinin kimyasal bileşimleri ve antimikrobiyal aktiviteleri. Mikrobiyol. Res. 160 , 189–195
Vandenberg, JD (1994) Farklı diyetlerle yetiştirilen yonca yaprağı kesen arının (Hymenoptera: Megachilidae) larvaları arasında tebeşir duyarlılığı. J. Econ. Entomol. 87 , 350–355
vanEngelsdorp, D., Evans, JD, Saegerman, C., Mullin, C., Haubruge, E., ve diğerleri. (2009) Koloni çöküşü bozukluğu: tanımlayıcı bir çalışma. PLoS ONE 4 , e6481
Vardar-Ünlü, G., Silici, S., Ünlü, M. (2008) Kavak tomurcukları ve kavak tipi propolisin bileşimi ve in vitro antimikrobiyal aktivitesi. Dünya J. Microbiol. Biyoteknoloji. 24 , 1011–1017
Vaudo, AD, Tooker, JF, Grozinger, CM, Patch, HM (2015) Arı beslenmesi ve çiçek kaynağı restorasyonu. Curr. Görüş. Böcek Bilimi. 10 , 133–141
Verge, J. (1951) Propolis, bal ve arı sütünün antibakteriyel aktivitesi. Arıcı 95 , 13–20
Visscher, P. (1980) Bal arılarının ( Apis mellifera ) yuva hijyeni sorunlarına adaptasyonu. Sosyobiyoloji 5 , 249–260
Walker, P., Crane, E. (1987) Propolisin bileşenleri. Apidologie 18 , 327–334
Wellford, TET, Eadie, T., Llewellyn, GC (1978) Balın mantar büyümesi, sporlanma ve aflatoksin üretimi üzerindeki önleyici etkisinin değerlendirilmesi. Z. Lebensm. Unters. Forsch. 166 , 280–283
White, GF (1919) Nosema hastalığı. ABD Tarım Bakanlığı Boğa. 780 , 1–59
Wilson, MB, Spivak, M., Hegeman, AD, Rendahl, A., Cohen, JD (2013) Metabolomik, bal arıları tarafından toplanan antimikrobiyal bitki reçinelerinin kökenlerini ortaya koyuyor. PLoS ONE 8 , e77512
Wilson, MB, Brinkman, D., Spivak, M., Gardner, G., Cohen, JD (2015) ABD propolisinin Paenibacillus larvaları ve Ascosphaera apis'ekarşı bileşimi ve antimikrobiyal aktivitesindeki bölgesel farklılıklar . J. Omurgasız. Pathol. 124 , 44–50
Wilson-Rich, N., Spivak, M., Fefferman, NH, Starks, PT (2009) Böcek toplumlarında hastalık direncinin genetik, bireysel ve grup olarak kolaylaştırılması. Annu. Rahip Entomol. 54 , 405–423
Zhang, Y., Zhang, G., Huang, X., Han, R. (2014) Heterospesifik arı sütü ve CSBV mücadelesi ile beslenen Apis cerana ve Apis mellifera larvalarının proteomik analizi . PLoS ONE 9 , e102663
- Cilt 47 , sayfalar 389–411, ( 2016 )
- Bu makaleden alıntı yap
Hiç yorum yok:
Yorum Gönder
Selam 🙋🏼♀️Hallo
*Hemsire (1-2-3-/∞) &Otodidaktik Araştırmaci Yazar.
Cahil bilmenin,Alim anlamanin pesindedir.-S.O